La membrana celular constituye una estructura dinámica, asimétrica y altamente especializada cuya función trasciende la simple delimitación física entre el medio intracelular y el extracelular. Su organización molecular permite mantener la homeostasis, regular el intercambio de sustancias, coordinar la comunicación intercelular y traducir señales químicas y mecánicas en respuestas biológicas específicas. Dentro de esta arquitectura, las proteínas de membrana representan los elementos funcionales más importantes, debido a que la bicapa lipídica por sí sola posee propiedades fisicoquímicas insuficientes para sostener la complejidad funcional requerida por las células vivas.
Las proteínas de membrana se clasifican en proteínas integrales y proteínas periféricas según su relación estructural con la bicapa lipídica. Ambas categorías participan de manera coordinada en procesos esenciales como transporte iónico, señalización celular, reconocimiento molecular, adhesión intercelular, transducción de señales, actividad enzimática y organización del citoesqueleto.
Las proteínas integrales de membrana se encuentran embebidas parcial o totalmente en la bicapa lipídica. Muchas de ellas atraviesan toda la membrana mediante dominios hidrofóbicos ricos en aminoácidos no polares que interactúan estrechamente con las colas lipídicas de los fosfolípidos. Esta disposición es energéticamente favorable porque minimiza el contacto entre regiones hidrofóbicas y el agua circundante. La mayor parte de estas proteínas son glucoproteínas, debido a que poseen cadenas de carbohidratos unidas covalentemente que protruyen hacia la superficie extracelular formando parte del glucocálix.
La glucosilación de las proteínas integrales posee enorme relevancia fisiológica. Los oligosacáridos participan en fenómenos de reconocimiento celular, interacción inmunológica, adhesión entre células y protección mecánica de la superficie celular. Además, las cadenas glucídicas contribuyen a estabilizar la conformación tridimensional de las proteínas y participan en mecanismos de clasificación intracelular durante el tráfico vesicular.
La razón por la que las proteínas integrales pueden “flotar” dentro de la membrana deriva del modelo de mosaico fluido. En este modelo, la bicapa lipídica actúa como una matriz fluida bidimensional en la que proteínas y lípidos poseen movilidad lateral relativa. Esta fluidez depende principalmente de la composición lipídica, del contenido de colesterol y del grado de saturación de los ácidos grasos. La movilidad proteica permite reorganizaciones dinámicas indispensables para señalización celular, formación de complejos multiproteicos, endocitosis y exocitosis.
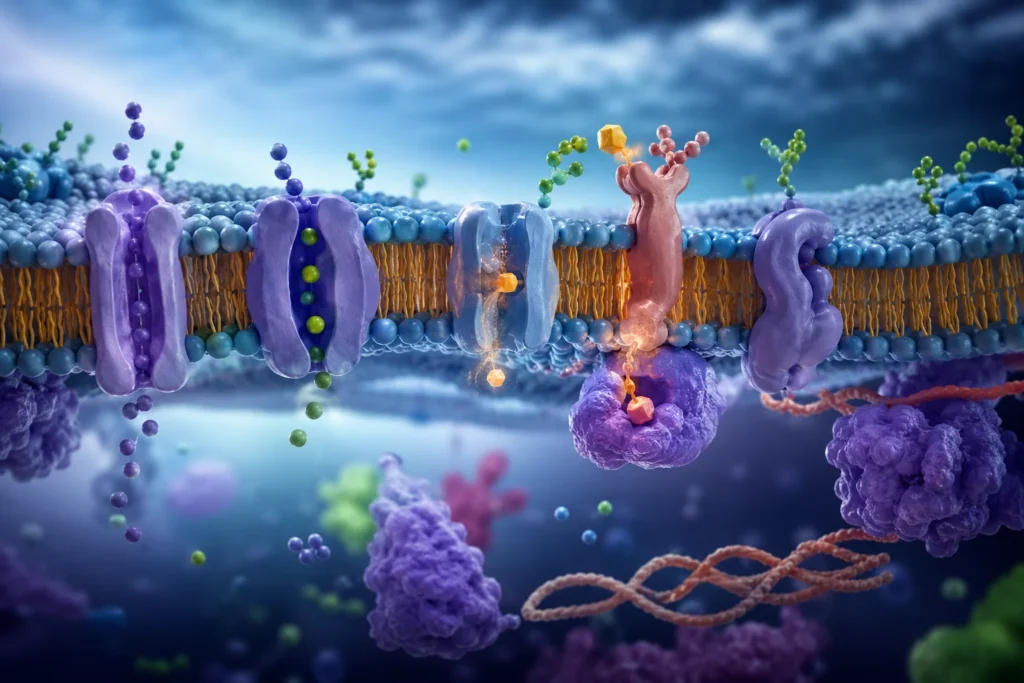
Muchas proteínas integrales funcionan como canales o poros transmembrana. Esta función es necesaria porque la bicapa lipídica posee un núcleo hidrofóbico que constituye una barrera energética extremadamente eficiente contra el paso espontáneo de moléculas polares y partículas cargadas. El agua puede difundir en pequeña proporción a través de la bicapa, pero el transporte rápido y regulado requiere proteínas especializadas llamadas acuaporinas.
Los canales iónicos poseen estructuras proteicas que generan un ambiente hidrofílico interno capaz de permitir el tránsito selectivo de iones específicos. La selectividad iónica depende de propiedades como diámetro del poro, distribución de cargas eléctricas y presencia de regiones denominadas filtros de selectividad. Por ejemplo, los canales de potasio permiten el paso eficiente de K+ mientras excluyen Na+, aun cuando este último posee menor radio atómico. Ello ocurre porque el filtro de selectividad estabiliza energéticamente al ion deshidratado de potasio mediante interacciones precisas entre oxígenos carbonilo y el radio iónico específico del K+.
La apertura y cierre de los canales iónicos constituye un fenómeno denominado compuerta o “gating”. Algunos canales son regulados por diferencias de voltaje, otros por unión de ligandos y otros por estímulos mecánicos. Esta regulación permite transformar estímulos físicos y químicos en señales electrofisiológicas. En neuronas y células musculares, los cambios rápidos de permeabilidad iónica generan potenciales de acción, contracción muscular y liberación de neurotransmisores.
Además de los canales, numerosas proteínas integrales actúan como transportadores o proteínas carrier. A diferencia de los canales, los transportadores se unen específicamente al soluto y experimentan cambios conformacionales alternantes que exponen el sitio de unión hacia uno u otro lado de la membrana. Este mecanismo permite el transporte selectivo de glucosa, aminoácidos, nucleósidos y múltiples metabolitos.
El transporte mediado puede ser pasivo o activo. En el transporte pasivo facilitado, el movimiento ocurre a favor del gradiente electroquímico y no requiere aporte directo de energía metabólica. Sin embargo, muchas sustancias deben desplazarse contra sus gradientes de concentración, lo que requiere transporte activo. Este proceso resulta fundamental para mantener diferencias iónicas esenciales para la excitabilidad celular, regulación osmótica y metabolismo.
Las proteínas responsables del transporte activo utilizan diversas fuentes de energía. El transporte activo primario depende directamente de la hidrólisis de ATP. Un ejemplo fundamental es la Na+/K+-ATPasa, que expulsa 3 iones Na+ e introduce 2 iones K+ por cada molécula de ATP hidrolizada. Esta actividad mantiene el potencial de membrana en reposo, regula el volumen celular y genera gradientes electroquímicos utilizados posteriormente para transporte secundario.
El transporte activo secundario utiliza la energía almacenada en gradientes previamente establecidos. En este mecanismo, el movimiento favorable de un ion impulsa el transporte desfavorable de otra sustancia. Un ejemplo clásico es el cotransportador Na+/glucosa del epitelio intestinal, mediante el cual el gradiente de sodio impulsa la absorción de glucosa incluso cuando la concentración intracelular de esta es elevada.
Otras proteínas integrales poseen actividad enzimática. Algunas participan directamente en cascadas metabólicas localizadas en la membrana, mientras otras funcionan como enzimas receptoras. La localización membranal permite integrar señales extracelulares con respuestas intracelulares inmediatas y espacialmente organizadas.
Las proteínas receptoras constituyen una de las funciones más sofisticadas de la membrana celular. Muchas moléculas señalizadoras son hidrosolubles y no pueden atravesar fácilmente la bicapa lipídica. Entre ellas se encuentran hormonas peptídicas, neurotransmisores y factores de crecimiento. Para transmitir información, estas moléculas se unen a receptores específicos localizados en la superficie celular.
La unión entre ligando y receptor desencadena cambios conformacionales altamente específicos. Dichas modificaciones estructurales alteran la actividad funcional del receptor y permiten la propagación de la señal hacia el interior celular. Este fenómeno representa la base molecular de la transducción de señales.
Algunos receptores poseen actividad enzimática intrínseca. Los receptores tirosina cinasa, por ejemplo, experimentan autofosforilación tras la unión del ligando. La fosforilación crea sitios de acoplamiento para proteínas intracelulares que activan cascadas de señalización relacionadas con proliferación celular, diferenciación, metabolismo y supervivencia.
Otros receptores se asocian a proteínas G heterotriméricas. La activación del receptor induce intercambio de GDP por GTP en la subunidad alfa de la proteína G, permitiendo la activación de enzimas efectoras como adenilato ciclasa o fosfolipasa C. Estas enzimas generan segundos mensajeros intracelulares como AMP cíclico, inositol trifosfato y diacilglicerol, moléculas capaces de amplificar extraordinariamente la señal original.
La amplificación constituye una propiedad esencial de la señalización celular. Una sola molécula hormonal puede activar miles de moléculas intracelulares en cascada, permitiendo respuestas biológicas intensas aun con concentraciones extracelulares mínimas del ligando.
La membrana celular funciona así como un sistema de percepción molecular extremadamente sofisticado. Las proteínas integrales no solo permiten intercambio de sustancias, sino que convierten información química, mecánica y eléctrica del entorno en modificaciones bioquímicas intracelulares precisas.
Por otro lado, las proteínas periféricas no atraviesan la bicapa lipídica. Se unen débilmente a la superficie interna o externa de la membrana mediante interacciones electrostáticas, puentes de hidrógeno o asociación con proteínas integrales. Debido a estas uniones relativamente débiles, pueden separarse sin alterar significativamente la estructura lipídica.
Las proteínas periféricas cumplen funciones regulatorias fundamentales. Muchas actúan como enzimas intracelulares asociadas a la membrana, donde organizan rutas metabólicas y cascadas de señalización. Otras participan en el anclaje del citoesqueleto a la membrana plasmática, proporcionando estabilidad mecánica, forma celular y coordinación del movimiento celular.
La interacción entre proteínas periféricas y proteínas integrales resulta esencial para el control funcional de canales y transportadores. Algunas proteínas periféricas modulan apertura de canales iónicos mediante fosforilación o interacciones conformacionales. Otras organizan complejos macromoleculares necesarios para señalización eficiente.
En eritrocitos, proteínas periféricas como espectrina, anquirina y proteína banda 4.1 forman una red submembranal que otorga elasticidad y resistencia mecánica a la célula. Alteraciones genéticas en estas proteínas producen enfermedades como esferocitosis hereditaria, caracterizada por fragilidad eritrocitaria y hemólisis.
Las proteínas periféricas también participan en fenómenos de tráfico vesicular. Complejos proteicos asociados transitoriamente a la membrana regulan formación, desplazamiento y fusión de vesículas intracelulares. Estos procesos son esenciales para secreción hormonal, neurotransmisión y reciclaje de receptores.
La organización funcional de proteínas integrales y periféricas no ocurre de manera aleatoria. La membrana posee microdominios especializados ricos en colesterol y esfingolípidos llamados balsas lipídicas. Estas regiones concentran proteínas específicas involucradas en señalización celular y tráfico membranal, aumentando la eficiencia funcional mediante proximidad molecular.
La alteración de proteínas de membrana tiene profundas consecuencias fisiopatológicas. Mutaciones en canales iónicos producen canalopatías responsables de epilepsia, arritmias cardíacas y miopatías. Alteraciones en receptores de membrana participan en diabetes mellitus, cáncer y enfermedades autoinmunes. Defectos en transportadores generan síndromes metabólicos, trastornos neurológicos y enfermedades renales.
La enorme diversidad funcional de las proteínas de membrana refleja la complejidad evolutiva de las células eucariotas. Estas proteínas constituyen sistemas moleculares capaces de integrar transporte, comunicación, reconocimiento y regulación metabólica dentro de una única plataforma estructural dinámica: la membrana celular.


Fuente y lecturas recomendadas:
- Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2022). Molecular Biology of the Cell (7th ed.). Garland Science.
- Cooper, G. M., & Hausman, R. E. (2019). The Cell: A Molecular Approach (8th ed.). Sinauer Associates.
- Guyton, A. C., & Hall, J. E. (2021). Textbook of Medical Physiology (14th ed.). Elsevier.
- Hille, B. (2001). Ion Channels of Excitable Membranes (3rd ed.). Sinauer Associates.
- Kandel, E. R., Koester, J. D., Mack, S. H., & Siegelbaum, S. A. (2021). Principles of Neural Science (6th ed.). McGraw-Hill.
- Lodish, H., Berk, A., Kaiser, C. A., Krieger, M., Bretscher, A., Ploegh, H., Amon, A., & Scott, M. P. (2021). Molecular Cell Biology (9th ed.). W. H. Freeman.
- Nicolson, G. L. (2014). The fluid-mosaic model of membrane structure: Still relevant to understanding the structure, function and dynamics of biological membranes after more than 40 years. Biochimica et Biophysica Acta – Biomembranes, 1838(6), 1451–1466.
- Singer, S. J., & Nicolson, G. L. (1972). The fluid mosaic model of the structure of cell membranes. Science, 175(4023), 720–731.
- Verkman, A. S. (2011). Aquaporins at a glance. Journal of Cell Science, 124(13), 2107–2112.
- Venter, J. C., Adams, M. D., Myers, E. W., Li, P. W., Mural, R. J., Sutton, G. G., Smith, H. O., Yandell, M., Evans, C. A., Holt, R. A., et al. (2001). The sequence of the human genome. Science, 291(5507), 1304–1351.
Aprende administración paso a paso

ADMINISTRACION DESDE CERO