La barrera lipídica de la membrana celular constituye uno de los principios fisicoquímicos más importantes de la biología celular, debido a que permite la existencia de un medio intracelular diferenciado del entorno extracelular. Esta propiedad no solamente delimita estructuralmente a la célula, sino que además hace posible la homeostasis, la excitabilidad eléctrica, la señalización molecular, el metabolismo energético, el transporte selectivo y la supervivencia celular. La membrana plasmática posee un grosor aproximado de 7,5 a 10 nm y está formada principalmente por lípidos y proteínas organizados en una disposición dinámica conocida como modelo del mosaico fluido. Su composición aproximada incluye un 55% de proteínas, un 25% de fosfolípidos, un 13% de colesterol, un 4% de otros lípidos y un 3% de hidratos de carbono. Esta organización molecular genera una barrera semipermeable altamente especializada capaz de discriminar entre moléculas según su tamaño, polaridad, carga eléctrica y solubilidad lipídica.
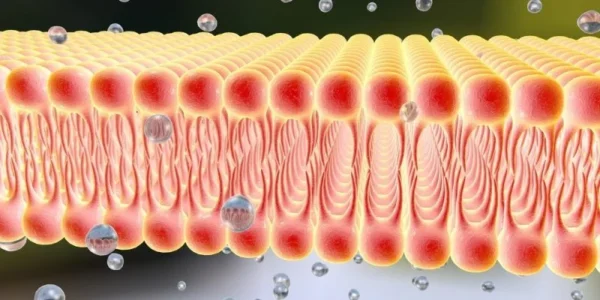
La razón fundamental por la cual la membrana celular actúa como una barrera frente a sustancias hidrosolubles radica en la naturaleza anfipática de los fosfolípidos que forman la bicapa lipídica. Los fosfolípidos contienen una cabeza polar hidrófila compuesta principalmente por grupos fosfato ionizados y dos colas hidrófobas formadas por cadenas hidrocarbonadas derivadas de ácidos grasos. Esta dualidad estructural determina espontáneamente su organización en medio acuoso. Cuando los fosfolípidos son expuestos al agua, las cabezas hidrófilas interactúan favorablemente con las moléculas polares del entorno mediante puentes de hidrógeno e interacciones electrostáticas, mientras que las colas hidrófobas son excluidas del contacto con el agua debido al efecto hidrofóbico. Como consecuencia, las moléculas se autoensamblan formando una bicapa en la cual las regiones hidrófobas quedan orientadas hacia el interior de la membrana y las regiones hidrófilas hacia los compartimentos acuosos intra y extracelulares. Este proceso ocurre de manera espontánea porque minimiza la energía libre del sistema y reduce el desorden inducido en las moléculas de agua circundantes.
La región central hidrófoba de la bicapa constituye la verdadera barrera de permeabilidad. Las moléculas hidrosolubles, especialmente aquellas que poseen carga eléctrica o elevada polaridad, presentan una cubierta de hidratación formada por moléculas de agua organizadas alrededor de ellas. Para atravesar la bicapa lipídica, dichas moléculas tendrían que desprenderse parcial o totalmente de esta capa de hidratación e ingresar en un entorno altamente apolar e hidrófobo. Este proceso requiere un gasto energético considerablemente elevado y resulta termodinámicamente desfavorable. Por esta razón, iones como sodio, potasio, calcio y cloruro poseen una permeabilidad extremadamente baja a través de la bicapa lipídica pura. Del mismo modo, moléculas polares relativamente grandes, como glucosa, aminoácidos y urea, atraviesan la membrana con enorme dificultad en ausencia de proteínas transportadoras específicas.
En contraste, las moléculas liposolubles atraviesan la membrana con relativa facilidad debido a que pueden disolverse transitoriamente en la región hidrófoba de la bicapa. Gases pequeños y no polares como oxígeno, dióxido de carbono y nitrógeno difunden rápidamente a través de la membrana siguiendo gradientes de concentración. De manera semejante, moléculas orgánicas hidrófobas como hormonas esteroideas, anestésicos liposolubles y ciertos alcoholes penetran con rapidez porque su estructura química es compatible con el entorno apolar de la bicapa. La velocidad de difusión depende de factores como el coeficiente de partición lípido-agua, el tamaño molecular, la temperatura y la composición específica de los lípidos membranales.
La impermeabilidad relativa de la bicapa lipídica frente a sustancias hidrosolubles tiene profundas implicaciones fisiológicas. En primer lugar, permite mantener gradientes electroquímicos entre el medio intracelular y extracelular. La concentración intracelular elevada de potasio y la concentración extracelular elevada de sodio son esenciales para la generación del potencial de membrana y la excitabilidad eléctrica de neuronas y fibras musculares. Si la bicapa lipídica fuese libremente permeable a estos iones, los gradientes iónicos se disiparían rápidamente y desaparecerían fenómenos fundamentales como el potencial de acción, la contracción muscular y la transmisión sináptica.
Además, la barrera lipídica permite conservar diferencias osmóticas y metabólicas entre el citoplasma y el exterior celular. El interior celular contiene elevadas concentraciones de proteínas, metabolitos y macromoléculas que deben permanecer confinados para garantizar la función enzimática, la síntesis proteica y el metabolismo energético. La pérdida descontrolada de estos componentes o la entrada indiscriminada de sustancias extracelulares comprometería rápidamente la viabilidad celular.
La bicapa lipídica no es una estructura rígida, sino una matriz fluida y dinámica. Las moléculas lipídicas poseen movilidad lateral constante dentro del plano de la membrana, fenómeno que permite redistribuir proteínas, fusionar membranas, formar vesículas y facilitar procesos de señalización celular. Esta fluidez depende de múltiples factores, especialmente de la composición de ácidos grasos y del contenido de colesterol. Los ácidos grasos insaturados contienen dobles enlaces que generan curvaturas en las cadenas hidrocarbonadas e impiden el empaquetamiento estrecho de los fosfolípidos. Como resultado, las membranas con mayor contenido de ácidos grasos insaturados presentan mayor fluidez y menor rigidez estructural.
El colesterol desempeña un papel esencial en la regulación de esta fluidez membranal. Debido a su estructura esteroidea rígida y planar, el colesterol se intercala entre las colas hidrófobas de los fosfolípidos y modula sus movimientos moleculares. A temperaturas elevadas, restringe el movimiento excesivo de los fosfolípidos y disminuye la fluidez; a temperaturas bajas, evita el empaquetamiento compacto de las cadenas hidrocarbonadas e impide la cristalización de la membrana. De esta manera, el colesterol actúa como un amortiguador de fluidez que estabiliza las propiedades físicas de la membrana frente a cambios térmicos.
Otra función fundamental del colesterol consiste en disminuir la permeabilidad de la bicapa a moléculas hidrosolubles pequeñas. El colesterol llena espacios intermoleculares existentes entre las cadenas de fosfolípidos y aumenta la compactación de la región hidrófoba central. Esto reduce el paso de agua y solutos polares a través de la membrana. La presencia de colesterol es especialmente importante en células animales, cuya supervivencia depende de una regulación precisa de la permeabilidad y estabilidad membranal.
Los esfingolípidos constituyen otro grupo importante de lípidos membranales. A diferencia de los fosfolípidos convencionales, derivan de la esfingosina y suelen contener cadenas hidrocarbonadas más largas y saturadas. Estas características favorecen interacciones intermoleculares intensas y generan regiones membranales más ordenadas y menos fluidas. Los esfingolípidos son particularmente abundantes en el tejido nervioso, especialmente en las vainas de mielina, donde contribuyen al aislamiento eléctrico de los axones y a la velocidad de conducción nerviosa.
Muchos esfingolípidos complejos participan también en señalización celular y reconocimiento molecular. Los glicoesfingolípidos poseen cadenas de hidratos de carbono orientadas hacia la superficie extracelular, donde funcionan como receptores, sitios de adhesión y marcadores de identidad celular. Algunas toxinas bacterianas, virus y microorganismos utilizan específicamente glicoesfingolípidos como puntos de unión para ingresar a las células. Asimismo, ciertos esfingolípidos participan en vías de señalización intracelular relacionadas con apoptosis, proliferación y diferenciación celular.
La membrana plasmática contiene microdominios especializados conocidos como balsas lipídicas, regiones enriquecidas en colesterol y esfingolípidos que presentan una organización más ordenada y menos fluida que el resto de la membrana. Estas estructuras concentran proteínas específicas involucradas en señalización celular, tráfico vesicular y organización del citoesqueleto. Las balsas lipídicas facilitan la interacción eficiente entre receptores, enzimas y moléculas señalizadoras, incrementando la velocidad y especificidad de numerosas respuestas celulares.
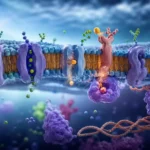
Aunque la bicapa lipídica constituye la base de la barrera de permeabilidad, las proteínas de membrana permiten el transporte selectivo de sustancias que no pueden atravesarla libremente. Existen proteínas canal que forman poros hidrofílicos para el paso rápido de iones y agua, así como proteínas transportadoras que cambian de conformación para movilizar moléculas específicas. Algunas proteínas funcionan mediante difusión facilitada y otras mediante transporte activo dependiente de energía metabólica. Gracias a esta combinación de impermeabilidad lipídica y permeabilidad proteica selectiva, la célula puede controlar rigurosamente el intercambio de sustancias con el entorno.
La importancia fisiopatológica de la barrera lipídica es enorme. Alteraciones en la composición lipídica de la membrana modifican la permeabilidad, la fluidez y la función celular. El estrés oxidativo puede inducir peroxidación lipídica, proceso en el cual radicales libres atacan los ácidos grasos insaturados de la membrana, generando pérdida de integridad estructural y aumento de la permeabilidad. Este fenómeno participa en envejecimiento celular, neurodegeneración, lesión isquémica y múltiples enfermedades inflamatorias.
Asimismo, defectos hereditarios en el metabolismo de esfingolípidos producen enfermedades de depósito lisosomal caracterizadas por acumulación patológica de lípidos en el sistema nervioso y otros tejidos. Alteraciones del colesterol membranal también afectan procesos de señalización, tráfico intracelular y susceptibilidad a infecciones virales.
La aparición de membranas lipídicas autoorganizadas representó un acontecimiento decisivo para el origen de la vida celular. La capacidad de separar un medio interno del ambiente externo permitió la concentración de biomoléculas, el establecimiento de gradientes energéticos y el desarrollo de sistemas metabólicos complejos. Sin la existencia de una barrera lipídica selectiva, la organización bioquímica necesaria para la vida sería imposible.


Fuente y lecturas recomendadas:
- Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2022). Molecular Biology of the Cell (7th ed.). Garland Science.
- Cooper, G. M., & Hausman, R. E. (2019). The Cell: A Molecular Approach (8th ed.). Sinauer Associates.
- Engelman, D. M. (2005). Membranes are more mosaic than fluid. Nature, 438(7068), 578–580.
- Lingwood, D., & Simons, K. (2010). Lipid rafts as a membrane-organizing principle. Science, 327(5961), 46–50.
- Nicolson, G. L. (2014). The Fluid-Mosaic Model of membrane structure: Still relevant to understanding the structure, function and dynamics of biological membranes after more than 40 years. Biochimica et Biophysica Acta, 1838(6), 1451–1466.
- Singer, S. J., & Nicolson, G. L. (1972). The fluid mosaic model of the structure of cell membranes. Science, 175(4023), 720–731.
- van Meer, G., Voelker, D. R., & Feigenson, G. W. (2008). Membrane lipids: Where they are and how they behave. Nature Reviews Molecular Cell Biology, 9(2), 112–124.
- Yeagle, P. L. (2016). The Membranes of Cells (3rd ed.). Academic Press.
- Zwiazek, J. J. (2022). Roles of membrane lipids in plant and animal cell physiology. Cells, 11(7), 1147.
Aprende administración paso a paso

ADMINISTRACION DESDE CERO