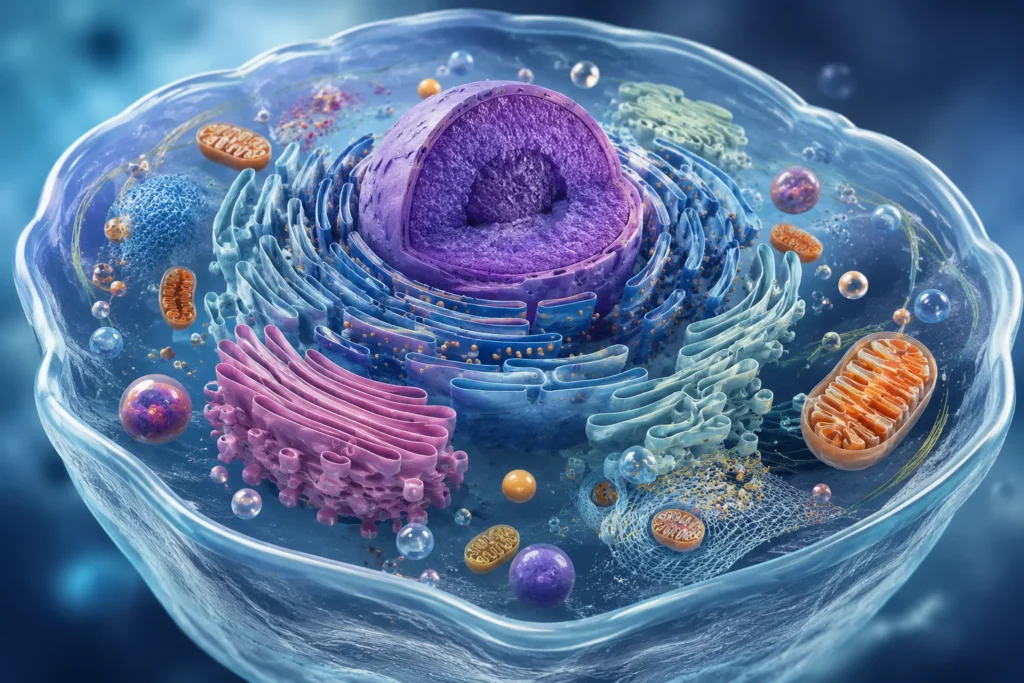
Las estructuras membranosas de la célula constituyen uno de los principios fundamentales de la organización biológica, debido a que permiten la compartimentalización del medio intracelular y la regulación precisa de prácticamente todos los procesos fisiológicos celulares. La célula eucariota no es simplemente una masa homogénea de sustancias químicas suspendidas en el citoplasma, sino un sistema altamente organizado en el que múltiples orgánulos delimitados por membranas cooperan de manera dinámica para mantener la homeostasis, la producción energética, la síntesis de biomoléculas, la degradación de sustancias, la comunicación intracelular y la señalización celular. Esta organización membranosa incrementa extraordinariamente la eficiencia metabólica porque permite que distintas reacciones químicas ocurran de manera simultánea en microambientes especializados con composiciones iónicas, concentraciones de sustratos y condiciones fisicoquímicas específicas.
Las membranas celulares están constituidas principalmente por una bicapa lipídica formada por fosfolípidos anfipáticos, colesterol, esfingolípidos y proteínas asociadas. La naturaleza anfipática de los fosfolípidos es esencial para la formación espontánea de la bicapa. Cada fosfolípido posee una región hidrofílica polar y una región hidrofóbica apolar. En presencia de agua, las regiones hidrofóbicas tienden a excluir el contacto con el medio acuoso, mientras que las regiones hidrofílicas interactúan con él. Como consecuencia de este fenómeno termodinámico, denominado efecto hidrofóbico, los fosfolípidos se autoorganizan espontáneamente formando bicapas estables en las que las colas hidrofóbicas quedan orientadas hacia el interior y las cabezas polares hacia el exterior. Este arreglo constituye la base estructural de todas las membranas biológicas.
La bicapa lipídica actúa como una barrera semipermeable que separa compartimientos acuosos con composiciones químicas diferentes. Esta propiedad es fundamental porque el interior hidrofóbico de la membrana impide el libre paso de moléculas hidrosolubles, iones y sustancias polares. El agua y las moléculas cargadas no pueden atravesar fácilmente la región hidrofóbica debido a incompatibilidades energéticas entre sus cargas eléctricas y las cadenas hidrocarbonadas apolares de los lípidos. Gracias a ello, la célula puede mantener gradientes electroquímicos, diferencias osmóticas y concentraciones específicas de metabolitos entre distintos compartimientos. Sin esta barrera selectiva sería imposible conservar la organización química necesaria para la vida celular.
La membrana plasmática constituye la estructura membranosa más externa de la célula y representa el límite físico entre el medio intracelular y el ambiente extracelular. Su función principal consiste en regular el intercambio selectivo de sustancias, recibir señales químicas, participar en la adhesión celular y mantener la integridad estructural de la célula. La membrana plasmática posee una organización dinámica descrita inicialmente mediante el modelo de mosaico fluido. Este modelo establece que las proteínas se encuentran insertadas o asociadas a una bicapa lipídica fluida, permitiendo movimientos laterales de los componentes membranosos. Sin embargo, investigaciones posteriores demostraron que las membranas poseen una organización mucho más compleja, con dominios lipídicos especializados, interacciones con el citoesqueleto y regiones funcionales altamente organizadas.
El colesterol desempeña una función esencial en la estabilidad y fluidez de las membranas. Este lípido se intercala entre las moléculas fosfolipídicas y modifica el empaquetamiento de la bicapa. A temperaturas elevadas disminuye la fluidez excesiva de la membrana, mientras que a temperaturas bajas evita la cristalización rígida de los fosfolípidos. Además, participa en la formación de microdominios especializados denominados balsas lipídicas, regiones enriquecidas en colesterol y esfingolípidos que organizan proteínas relacionadas con señalización celular, tráfico vesicular y reconocimiento celular.
Las proteínas de membrana son indispensables para las funciones celulares porque la bicapa lipídica por sí sola constituye una barrera prácticamente impermeable para muchas sustancias necesarias para la vida celular. Existen proteínas integrales, que atraviesan completamente la membrana, y proteínas periféricas, que se asocian de manera más superficial mediante interacciones electrostáticas o con otras proteínas. Las proteínas integrales contienen regiones hidrofóbicas que interactúan con el núcleo lipídico de la bicapa y regiones hidrofílicas orientadas hacia los medios acuosos intra y extracelulares. Esta organización estructural les permite funcionar como canales, transportadores, receptores, enzimas y moléculas de adhesión.
Los canales proteicos permiten el paso selectivo de iones o moléculas pequeñas mediante poros hidrofílicos que atraviesan la membrana. La selectividad de estos canales depende del diámetro del poro, la distribución de cargas eléctricas y la conformación tridimensional de las proteínas. Gracias a estos mecanismos la célula puede regular con enorme precisión el movimiento de sodio, potasio, calcio, cloro y otros iones fundamentales para la excitabilidad eléctrica, la contracción muscular, la secreción y la transmisión sináptica.
Otras proteínas membranosas funcionan como transportadores activos o pasivos. Los transportadores pasivos facilitan el movimiento de sustancias a favor de gradientes de concentración, mientras que los transportadores activos utilizan energía, generalmente derivada de la hidrólisis de ATP, para movilizar sustancias contra gradientes electroquímicos. Este transporte activo resulta indispensable para mantener diferencias iónicas entre el interior y el exterior celular, conservar el potencial de membrana y permitir procesos como absorción intestinal, secreción glandular y transmisión nerviosa.
Muchas proteínas membranosas poseen actividad enzimática. Estas enzimas catalizan reacciones químicas localizadas directamente sobre las membranas, permitiendo que múltiples procesos metabólicos ocurran con alta eficiencia espacial. Por ejemplo, enzimas localizadas en la membrana mitocondrial interna participan en la cadena de transporte de electrones y la fosforilación oxidativa, mientras que enzimas del retículo endoplásmico intervienen en síntesis lipídica, detoxificación y modificaciones postraduccionales de proteínas. La asociación de enzimas a membranas facilita la concentración local de sustratos y la coordinación secuencial de reacciones metabólicas complejas.
La membrana nuclear rodea el núcleo celular y separa el material genético del citoplasma. Esta estructura posee una doble membrana con poros nucleares especializados que regulan el intercambio selectivo de ARN, proteínas y otras macromoléculas entre núcleo y citoplasma. La compartimentalización nuclear protege el ADN y permite una regulación precisa de replicación, transcripción y reparación genética. Además, la membrana nuclear se encuentra conectada estructuralmente con el retículo endoplásmico, estableciendo continuidad funcional entre ambos sistemas membranosos.
El retículo endoplásmico constituye una extensa red membranosa intracelular dividida en retículo endoplásmico rugoso y retículo endoplásmico liso. El retículo rugoso posee ribosomas adheridos a su superficie citoplasmática y participa principalmente en síntesis, plegamiento y modificación inicial de proteínas destinadas a secreción, membranas o lisosomas. El retículo liso participa en síntesis de lípidos, metabolismo de carbohidratos, detoxificación de sustancias y almacenamiento de calcio intracelular. La extensa superficie membranosa del retículo incrementa enormemente la capacidad metabólica celular.
El aparato de Golgi está constituido por cisternas membranosas apiladas que modifican, clasifican y distribuyen proteínas y lípidos sintetizados en el retículo endoplásmico. En este orgánulo ocurren glucosilaciones, fosforilaciones y empaquetamiento de moléculas destinadas a secreción, membranas celulares o lisosomas. La compartimentalización interna del aparato de Golgi permite una secuencia ordenada de modificaciones bioquímicas altamente específicas.
Las mitocondrias poseen dos membranas especializadas con funciones distintas. La membrana externa contiene proteínas porinas relativamente permeables, mientras que la membrana interna es extraordinariamente selectiva y contiene complejos proteicos de la cadena respiratoria. La membrana interna forma crestas mitocondriales que aumentan el área superficial disponible para fosforilación oxidativa y síntesis de ATP. Esta organización membranosa es esencial para mantener gradientes de protones utilizados en producción energética celular.
Los lisosomas son orgánulos delimitados por membranas que contienen enzimas hidrolíticas capaces de degradar proteínas, lípidos, carbohidratos y ácidos nucleicos. La membrana lisosomal protege al citoplasma de la acción destructiva de estas enzimas y mantiene un medio ácido necesario para su funcionamiento. Sin esta compartimentalización, las hidrolasas lisosomales podrían provocar autodigestión celular masiva.
Las membranas celulares no son estructuras estáticas. Poseen una dinámica compleja influenciada por interacciones entre lípidos, proteínas, citoesqueleto y matriz extracelular. El citoesqueleto restringe y organiza el movimiento lateral de proteínas membranosas formando dominios funcionales especializados. Esta organización dinámica permite respuestas rápidas a estímulos mecánicos, químicos y eléctricos. Además, la movilidad de proteínas y lípidos facilita procesos como endocitosis, exocitosis, migración celular y formación de contactos intercelulares.
La asimetría membranosa constituye otra propiedad fundamental. Las dos capas de la bicapa lipídica poseen composiciones diferentes de fosfolípidos y proteínas. Esta distribución asimétrica es esencial para señalización celular, reconocimiento inmunológico, coagulación y apoptosis. El mantenimiento de esta asimetría requiere enzimas especializadas que transportan selectivamente lípidos entre ambas monocapas membranosas.
Las estructuras membranosas permiten que la célula mantenga una organización espacial altamente especializada. Gracias a la compartimentalización membranosa pueden coexistir simultáneamente procesos metabólicos incompatibles, generarse gradientes energéticos, regularse intercambios moleculares y coordinarse respuestas celulares complejas. Las membranas representan, por tanto, no solo barreras físicas, sino plataformas dinámicas de integración bioquímica, señalización molecular y regulación fisiológica indispensables para la vida celular.


Fuente y lecturas recomendadas:
- Nicolson, G. L. (2013). Update of the 1972 Singer-Nicolson Fluid-Mosaic Model of Membrane Structure. Discoveries, 1(1), e3. https://doi.org/10.15190/d.2013.3
- Nicolson, G. L., & Ferreira de Mattos, G. (2021). A Brief Introduction to Some Aspects of the Fluid-Mosaic Model of Cell Membrane Structure and Its Importance in Membrane Lipid Replacement. Membranes, 11(12), 947. https://doi.org/10.3390/membranes11120947
- Nicolson, G. L. (2015). Cell Membrane Fluid–Mosaic Structure and Cancer Metastasis. Cancer Research, 75(7), 1169-1176. https://doi.org/10.1158/0008-5472.CAN-14-3216
- Nicolson, G. L., & Ferreira de Mattos, G. (2022). Fifty Years of the Fluid-Mosaic Model of Biomembrane Structure and Organization and Its Importance in Biomedicine with Particular Emphasis on Membrane Lipid Replacement. Biomedicines, 10(7), 1711. https://doi.org/10.3390/biomedicines10071711
- Mayor, S., et al. (2023). A Survey of Models of Cell Membranes: Toward a New Understanding of Membrane Organization. Cold Spring Harbor Perspectives in Biology, 15(9), a041976. https://doi.org/10.1101/cshperspect.a041976
Aprende administración paso a paso

ADMINISTRACION DESDE CERO