El ARNmi (microARN) y el ARN de interferencia pequeño constituyen dos de los principales mecanismos de regulación génica postranscripcional identificados en organismos eucariotas. Su importancia biológica radica en que permiten modular con gran precisión la cantidad de proteínas sintetizadas por una célula mediante la regulación de la estabilidad y traducción de los ARN mensajeros. Estos mecanismos representan una capa reguladora adicional a la regulación transcripcional clásica y participan en prácticamente todos los procesos celulares, incluyendo proliferación, diferenciación, apoptosis, desarrollo embrionario, homeostasis tisular, respuesta inmunitaria y adaptación a cambios ambientales.
La relevancia de estas moléculas quedó demostrada cuando se observó que pequeños ARN de doble cadena eran capaces de inducir un silenciamiento génico altamente específico. Este fenómeno, denominado interferencia por ARN, reveló la existencia de sistemas celulares conservados evolutivamente capaces de utilizar pequeñas secuencias de ARN como guías moleculares para identificar y regular ARN complementarios.
Los ARNmi son pequeñas moléculas monocatenarias de ARN no codificante compuestas generalmente por entre 21 y 23 nucleótidos. Se denominan ARN no codificantes porque no contienen la información necesaria para producir proteínas, sino que ejercen funciones regulatorias directas sobre la expresión génica.
Su descubrimiento modificó profundamente la comprensión de la genética molecular debido a que durante décadas se asumió que la función principal del ARN consistía en servir como intermediario entre ADN y proteínas. Posteriormente se comprobó que una proporción importante del genoma produce ARN funcionales que nunca son traducidos, pero que desempeñan funciones regulatorias esenciales.
Los genes que codifican ARNmi se encuentran distribuidos a lo largo del genoma. Algunos se localizan en regiones intergénicas independientes, mientras que otros están contenidos dentro de intrones o incluso exones de genes codificantes. La transcripción de estos genes es realizada principalmente por la ARN polimerasa II, generando transcritos primarios denominados ARNmi-pri.
Estos transcritos primarios pueden alcanzar cientos o miles de nucleótidos de longitud y presentan regiones complementarias que se aparean formando estructuras secundarias en horquilla. Dichas estructuras constituyen la señal molecular reconocida por la maquinaria celular encargada del procesamiento de los ARNmi.
La existencia de genes específicos para ARNmi demuestra que estas moléculas no son simples productos accidentales del metabolismo celular, sino elementos reguladores codificados y seleccionados evolutivamente para controlar redes complejas de expresión génica.
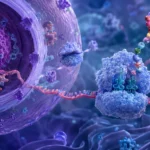
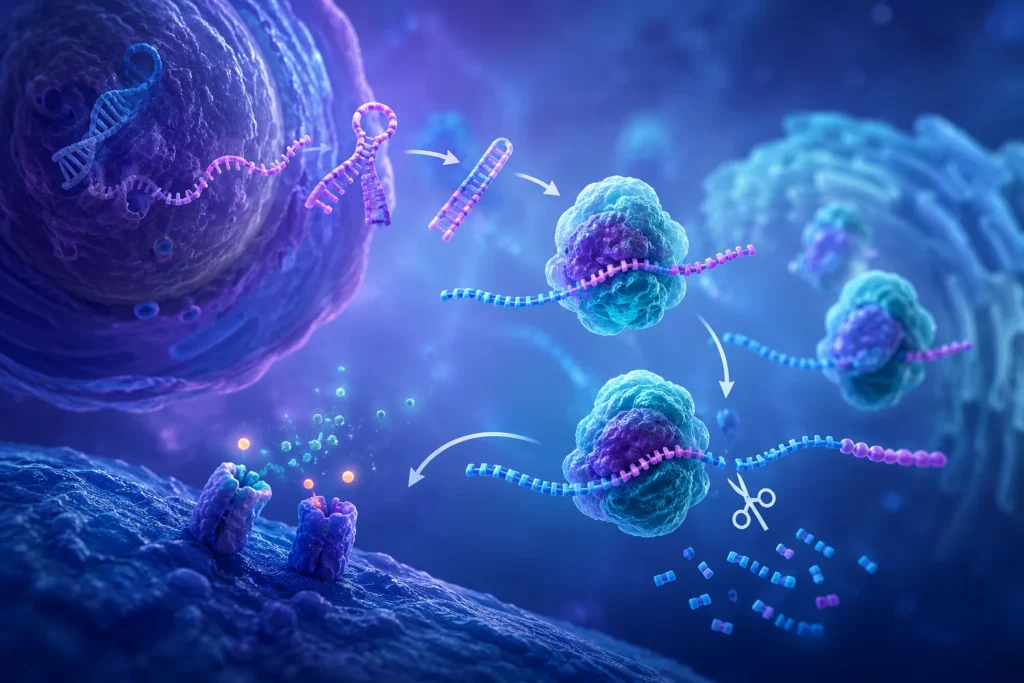
La generación de ARNmi maduros requiere una serie de eventos secuenciales altamente regulados que ocurren tanto en el núcleo como en el citoplasma.
La primera etapa consiste en la transcripción del gen correspondiente, originando el ARNmi-pri. Estos transcritos contienen una o varias estructuras de horquilla que serán procesadas posteriormente.
En el núcleo celular, el ARNmi-pri es reconocido por el complejo microprocesador, constituido principalmente por la ribonucleasa III Drosha y la proteína DGCR8. Este complejo identifica la estructura de horquilla y realiza un corte preciso que libera un precursor denominado pre-ARNmi.
El pre-ARNmi posee aproximadamente 70 nucleótidos y conserva una estructura característica en forma de horquilla. Posteriormente es transportado desde el núcleo hacia el citoplasma mediante la proteína Exportin-5, dependiente de Ran-GTP.
Una vez en el citoplasma, el pre-ARNmi es reconocido por otra ribonucleasa III denominada Dicer. Esta enzima elimina el bucle terminal de la horquilla y genera un pequeño dúplex de ARN de aproximadamente 22 nucleótidos.
Después del procesamiento por Dicer, una de las cadenas del dúplex es seleccionada como cadena guía y se incorpora al complejo de silenciamiento inducido por ARN. La cadena complementaria, denominada cadena pasajera, suele degradarse.
La incorporación de la cadena guía genera el ARNmi funcional maduro, capaz de reconocer secuencias específicas en ARN mensajeros diana y regular su expresión.
La extraordinaria precisión de este procesamiento es fundamental para la función biológica de los ARNmi, ya que incluso pequeñas alteraciones en la secuencia pueden modificar profundamente el conjunto de genes regulados.
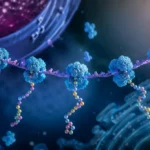
El complejo de silenciamiento inducido por ARN constituye la maquinaria efectora central de los mecanismos de interferencia por ARN.
Este complejo está formado por proteínas especializadas, entre las cuales destacan las proteínas Argonauta. Estas proteínas actúan como plataformas moleculares capaces de unir los pequeños ARN reguladores y utilizarlos como guías para localizar moléculas complementarias.
La secuencia del ARNmi dirige el complejo hacia ARN mensajeros específicos mediante complementariedad de bases. Una región particularmente importante es la denominada región semilla, ubicada generalmente entre los nucleótidos 2 y 8 del ARNmi.
Debido a que una misma secuencia semilla puede encontrarse en numerosos ARN mensajeros, un único ARNmi tiene capacidad para regular simultáneamente decenas o incluso centenares de genes distintos.
Esta característica convierte a los ARNmi en reguladores maestros capaces de coordinar programas celulares completos en lugar de actuar únicamente sobre genes individuales.
Los ARNmi regulan la expresión génica principalmente mediante dos mecanismos. El primero consiste en la inhibición de la traducción. Cuando el ARNmi se une parcialmente a su ARN mensajero diana, puede impedir que los ribosomas sinteticen la proteína correspondiente.
El segundo mecanismo consiste en promover la degradación del ARN mensajero. La unión del ARNmi favorece el reclutamiento de complejos celulares que eliminan la cola poliadenilada, inducen descapuchamiento y finalmente degradan la molécula.
En animales, la complementariedad entre ARNmi y ARN mensajero suele ser imperfecta, por lo que predomina una combinación de represión traduccional y degradación progresiva del ARN.
Como consecuencia, la cantidad de proteína sintetizada disminuye significativamente, permitiendo a la célula ajustar con precisión los niveles de expresión génica.
Este sistema proporciona una regulación extremadamente flexible y dinámica, capaz de responder rápidamente a cambios fisiológicos o ambientales.

Funciones fisiológicas de los ARNmi
Los ARNmi participan en prácticamente todos los aspectos de la biología celular. Durante el desarrollo embrionario regulan la especificación celular, la formación de tejidos y la diferenciación de órganos. La eliminación experimental de componentes esenciales de la maquinaria de ARNmi produce alteraciones severas del desarrollo e incluso letalidad embrionaria.
En tejidos adultos controlan la proliferación celular, la supervivencia, la apoptosis y la renovación tisular. También participan en la regulación del metabolismo energético, la señalización hormonal y la respuesta frente al estrés celular.
En el sistema inmunitario intervienen en la diferenciación y activación de linfocitos, macrófagos y otras células inmunes, modulando tanto la inmunidad innata como la adaptativa.
En el sistema cardiovascular regulan procesos relacionados con hipertrofia cardíaca, angiogénesis, remodelado vascular y respuesta al daño tisular.
En el sistema nervioso participan en neurogénesis, plasticidad sináptica, aprendizaje y memoria. La amplitud de estas funciones refleja que los ARNmi forman parte integral de las redes reguladoras que mantienen la homeostasis celular.
Debido a que los ARNmi controlan numerosos genes simultáneamente, las alteraciones en su expresión pueden producir consecuencias patológicas importantes.
En cáncer se han identificado perfiles específicos de expresión de ARNmi asociados con distintos tipos tumorales. Algunos ARNmi actúan como supresores tumorales inhibiendo oncogenes, mientras que otros funcionan como oncogenes reguladores favoreciendo proliferación, invasión, angiogénesis y metástasis.
Alteraciones en ARNmi específicos también se han asociado con enfermedades cardiovasculares, incluyendo insuficiencia cardíaca, hipertensión arterial, aterosclerosis e infarto de miocardio.
En enfermedades neurodegenerativas se han descrito modificaciones de ARNmi relacionadas con procesos patológicos observados en enfermedad de Alzheimer, enfermedad de Parkinson y esclerosis lateral amiotrófica.
Asimismo, múltiples estudios han vinculado alteraciones de ARNmi con diabetes mellitus, enfermedades autoinmunes, fibrosis y diversas enfermedades infecciosas.
Estas observaciones han impulsado el desarrollo de ARNmi como biomarcadores diagnósticos y pronósticos.
ARN de interferencia pequeño
El ARN de interferencia pequeño es una molécula de ARN bicatenario de aproximadamente 20 a 25 nucleótidos que participa en el silenciamiento génico mediante mecanismos relacionados con los ARNmi.
Aunque ambos tipos de ARN utilizan el complejo de silenciamiento inducido por ARN y comparten componentes de procesamiento, existen diferencias fundamentales en su origen y modo de acción.
Los ARNmi se originan a partir de genes endógenos específicos y suelen presentar complementariedad parcial con sus ARN mensajeros diana. Por el contrario, los ARN de interferencia pequeños suelen derivar de ARN bicatenarios largos, ya sean exógenos o sintéticos, y generalmente presentan complementariedad casi perfecta con sus secuencias objetivo.
Esta diferencia determina importantes consecuencias funcionales.
Los ARN de interferencia pequeños pueden originarse a partir de ARN bicatenarios introducidos experimentalmente, ARN virales o moléculas endógenas que generan estructuras de doble cadena.
Estos ARN bicatenarios son reconocidos directamente por Dicer en el citoplasma.
La enzima corta el ARN precursor produciendo pequeños fragmentos de aproximadamente 21 a 23 nucleótidos con extremos característicos.
Posteriormente, estos fragmentos se incorporan al complejo de silenciamiento inducido por ARN.
Al igual que ocurre con los ARNmi, una de las cadenas es seleccionada como guía, mientras que la otra es eliminada.
La cadena guía dirige entonces el complejo hacia ARN mensajeros complementarios.
La elevada complementariedad entre el ARN de interferencia pequeño y su ARN mensajero objetivo permite que las proteínas Argonauta ejerzan actividad endonucleasa.
Como resultado, el ARN mensajero es cortado en una posición específica y posteriormente degradado por otras nucleasas celulares.
Este mecanismo suele producir un silenciamiento génico más potente y específico que el generado por muchos ARNmi naturales.
Debido a que la degradación ocurre antes de que la traducción pueda completarse, la síntesis proteica correspondiente disminuye drásticamente.
Por esta razón, los ARN de interferencia pequeños constituyen herramientas extremadamente eficaces para estudiar la función génica.
La posibilidad de diseñar ARN de interferencia pequeños complementarios a prácticamente cualquier secuencia conocida ha revolucionado la investigación biomédica.
En estudios experimentales permiten reducir selectivamente la expresión de genes específicos para analizar sus funciones biológicas.
Esta estrategia ha facilitado la identificación de numerosas rutas moleculares implicadas en desarrollo, cáncer, metabolismo, inmunidad y neurobiología.
Desde el punto de vista terapéutico, los ARN de interferencia pequeños ofrecen la posibilidad de silenciar genes causantes de enfermedad sin modificar permanentemente el ADN.
El desarrollo tecnológico de sistemas de administración, nanopartículas lipídicas y conjugados moleculares ha permitido trasladar esta estrategia a la práctica clínica.
La aprobación de tratamientos basados en ARN de interferencia pequeño para enfermedades hereditarias ha demostrado que la interferencia por ARN puede utilizarse de manera segura y eficaz en humanos.
Entre las ventajas potenciales destacan la elevada especificidad, la capacidad de dirigirse a genes previamente considerados intratables y la posibilidad de diseñar nuevos tratamientos con relativa rapidez una vez conocida la secuencia génica implicada.
Comparación entre ARNmi y ARN de interferencia pequeño
Aunque ambos utilizan mecanismos de interferencia por ARN, presentan diferencias fundamentales.
Los ARNmi son reguladores fisiológicos endógenos producidos por genes específicos del organismo. Suelen reconocer múltiples ARN mensajeros mediante complementariedad parcial y generan una regulación fina de redes génicas complejas.
Los ARN de interferencia pequeños, en cambio, suelen actuar mediante complementariedad casi perfecta y provocan degradación directa del ARN mensajero objetivo.
Los ARNmi participan principalmente en la regulación fisiológica normal de la expresión génica, mientras que los ARN de interferencia pequeños se han convertido en herramientas experimentales y terapéuticas especialmente útiles para el silenciamiento dirigido de genes concretos.
Ambos sistemas constituyen ejemplos extraordinarios de cómo pequeñas moléculas de ARN pueden controlar procesos biológicos complejos y representan uno de los avances conceptuales más importantes de la biología molecular moderna.

Fuente y lecturas recomendadas:
- Bartel, D. P. (2004). MicroRNAs: Genomics, biogenesis, mechanism, and function. Cell, 116(2), 281–297. https://doi.org/10.1016/S0092-8674(04)00045-5
- Bernstein, E., Caudy, A. A., Hammond, S. M., & Hannon, G. J. (2001). Role for a bidentate ribonuclease in the initiation step of RNA interference. Nature, 409(6818), 363–366. https://doi.org/10.1038/35053110
- Bushati, N., & Cohen, S. M. (2007). MicroRNA functions. Annual Review of Cell and Developmental Biology, 23, 175–205. https://doi.org/10.1146/annurev.cellbio.23.090506.123406
- Fire, A., Xu, S., Montgomery, M. K., Kostas, S. A., Driver, S. E., & Mello, C. C. (1998). Potent and specific genetic interference by double-stranded RNA in Caenorhabditis elegans. Nature, 391(6669), 806–811. https://doi.org/10.1038/35888
- Ha, M., & Kim, V. N. (2014). Regulation of microRNA biogenesis. Nature Reviews Molecular Cell Biology, 15(8), 509–524. https://doi.org/10.1038/nrm3838
- Hammond, S. M. (2005). Dicing and slicing: The core machinery of the RNA interference pathway. FEBS Letters, 579(26), 5822–5829. https://doi.org/10.1016/j.febslet.2005.08.057
- He, L., & Hannon, G. J. (2004). MicroRNAs: Small RNAs with a big role in gene regulation. Nature Reviews Genetics, 5(7), 522–531. https://doi.org/10.1038/nrg1379
- Kim, V. N., Han, J., & Siomi, M. C. (2009). Biogenesis of small RNAs in animals. Nature Reviews Molecular Cell Biology, 10(2), 126–139. https://doi.org/10.1038/nrm2632
- Meister, G., & Tuschl, T. (2004). Mechanisms of gene silencing by double-stranded RNA. Nature, 431(7006), 343–349. https://doi.org/10.1038/nature02873
- Rana, T. M. (2007). Illuminating the silence: Understanding the structure and function of small RNAs. Nature Reviews Molecular Cell Biology, 8(1), 23–36. https://doi.org/10.1038/nrm2085
- Wilson, R. C., & Doudna, J. A. (2013). Molecular mechanisms of RNA interference. Annual Review of Biophysics, 42, 217–239. https://doi.org/10.1146/annurev-biophys-083012-130404
- Zamore, P. D., Tuschl, T., Sharp, P. A., & Bartel, D. P. (2000). RNAi: Double-stranded RNA directs the ATP-dependent cleavage of mRNA at 21 to 23 nucleotide intervals. Cell, 101(1), 25–33. https://doi.org/10.1016/S0092-8674(00)80620-0
Aprende administración paso a paso

ADMINISTRACION DESDE CERO