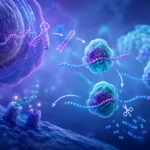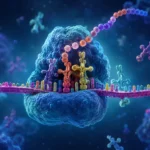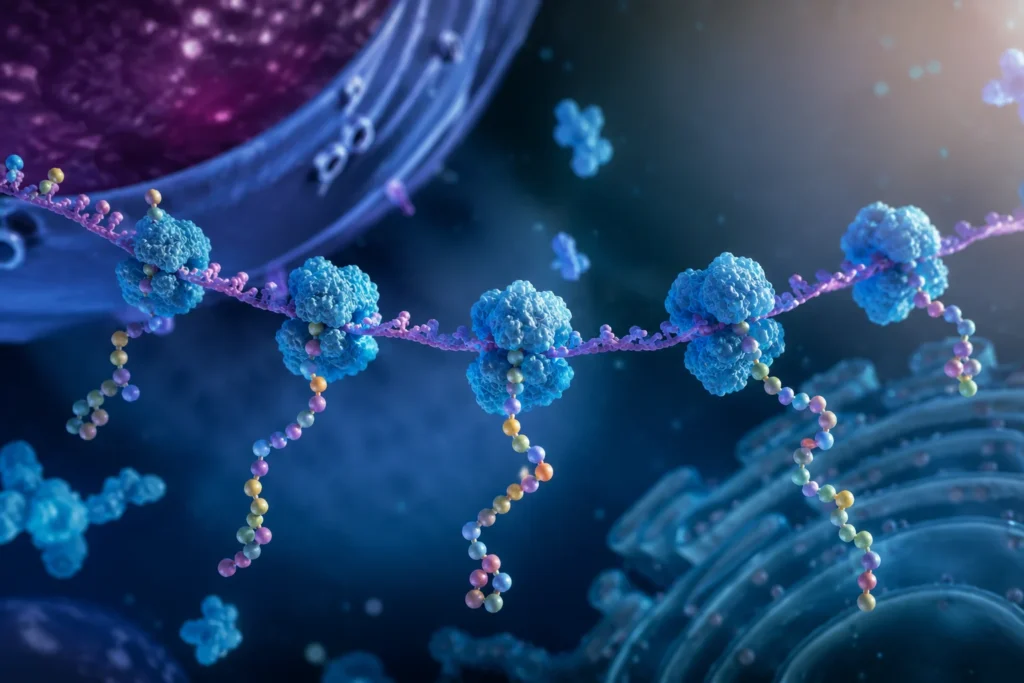
Los polirribosomas, también denominados polisomas, constituyen una de las estrategias más eficientes desarrolladas por las células para maximizar la producción de proteínas a partir de una única molécula de ARN mensajero. Este mecanismo permite que múltiples ribosomas traduzcan simultáneamente la misma secuencia de ARN mensajero, incrementando de manera extraordinaria la velocidad de síntesis proteica y optimizando el aprovechamiento de la información genética contenida en cada transcrito.
La base molecular de este fenómeno reside en la naturaleza secuencial de la traducción. Cuando un ARN mensajero abandona el núcleo en las células eucariotas o es sintetizado en el citoplasma en las células procariotas, se convierte en una plantilla accesible para los ribosomas. El primer ribosoma reconoce la región de inicio de la traducción y comienza a desplazarse a lo largo de la molécula de ARN mensajero en dirección 5’→3’, incorporando aminoácidos de acuerdo con la secuencia de codones presente en el transcrito. Durante este proceso, el extremo inicial del ARN mensajero queda nuevamente libre después de que el ribosoma avanza una distancia suficiente. Esta circunstancia permite que un segundo ribosoma se una al mismo ARN mensajero e inicie otra ronda de traducción antes de que el primer ribosoma haya completado la síntesis proteica. El mismo fenómeno puede repetirse numerosas veces, generando una fila de ribosomas distribuidos a lo largo de la molécula de ARN mensajero.
Como resultado, una sola molécula de ARN mensajero puede estar siendo leída simultáneamente por varios ribosomas, cada uno sintetizando una copia independiente de la misma proteína. Esta organización incrementa notablemente la eficiencia celular, ya que evita la necesidad de producir grandes cantidades adicionales de ARN mensajero para obtener múltiples moléculas proteicas. La formación de polisomas representa, por tanto, una estrategia fundamental de amplificación de la expresión génica.
La existencia de polirribosomas responde a principios termodinámicos y económicos de la biología celular. La síntesis de ARN mensajero requiere una considerable inversión energética, incluyendo la apertura de la cromatina, la actividad de la ARN polimerasa, el procesamiento postranscripcional y el transporte celular. Debido a este elevado costo metabólico, resulta más eficiente utilizar repetidamente la misma molécula de ARN mensajero mediante la unión simultánea de numerosos ribosomas. De esta manera, la célula obtiene una gran cantidad de proteína a partir de una única inversión inicial en transcripción.
La organización espacial de los polisomas presenta características particulares. Los ribosomas no se distribuyen aleatoriamente sobre el ARN mensajero, sino que mantienen una separación mínima determinada por sus dimensiones físicas y por la velocidad de elongación de la traducción. Un ribosoma ocupa aproximadamente entre 25 y 35 nucleótidos del ARN mensajero, por lo que los complejos de traducción sucesivos deben conservar una distancia suficiente para evitar colisiones estéricas. La cantidad exacta de ribosomas asociados a un mismo ARN mensajero depende de múltiples factores, incluyendo la longitud del transcrito, la eficiencia de iniciación, la velocidad de elongación y las condiciones fisiológicas de la célula.
Aunque tradicionalmente se describen polisomas formados por entre 3 y 10 ribosomas, numerosos estudios han demostrado que los ARN mensajeros largos pueden albergar decenas de ribosomas simultáneamente. En células altamente activas desde el punto de vista biosintético, algunos transcritos presentan densidades de traducción excepcionalmente elevadas, permitiendo la producción masiva de proteínas en intervalos muy breves.
Cada ribosoma presente en un polisoma se encuentra generalmente en una etapa diferente de la síntesis proteica. Los ribosomas más cercanos al extremo 5’ del ARN mensajero suelen contener cadenas polipeptídicas cortas, ya que han iniciado recientemente la traducción. En contraste, los ribosomas situados hacia el extremo 3’ contienen cadenas más largas y próximas a completarse. Esta disposición genera una especie de gradiente de crecimiento proteico a lo largo de toda la molécula de ARN mensajero.
Las observaciones realizadas mediante microscopía electrónica han revelado estructuras con apariencia de cuentas unidas por un hilo, donde el hilo corresponde al ARN mensajero y las cuentas representan los ribosomas individuales. Dependiendo de las características del transcrito y de las condiciones celulares, los polisomas pueden adoptar configuraciones lineales, circulares, helicoidales o compactadas.
En células eucariotas, la circularización funcional del ARN mensajero desempeña un papel importante en la formación y estabilidad de los polisomas. El extremo 5’ del ARN mensajero contiene una estructura denominada cap, mientras que el extremo 3’ posee una cola de poliadenina. Diversas proteínas establecen interacciones entre ambos extremos, formando un complejo cerrado que favorece el reciclaje de los ribosomas. Cuando un ribosoma completa la traducción y alcanza el codón de terminación, puede ser reutilizado eficientemente para iniciar una nueva ronda de síntesis sobre la misma molécula de ARN mensajero. Este mecanismo incrementa aún más la productividad translacional.
Los polisomas constituyen además un importante punto de regulación de la expresión génica. La cantidad de ribosomas asociados a un ARN mensajero determinado refleja directamente el nivel de traducción de ese gen. Los transcritos altamente expresados suelen encontrarse asociados con numerosos ribosomas, mientras que los ARN mensajeros poco activos presentan pocos ribosomas o incluso permanecen temporalmente almacenados sin traducirse. Por esta razón, el análisis de perfiles polisomales se ha convertido en una herramienta fundamental para estudiar la regulación génica a nivel translacional.
La formación de polisomas también permite respuestas celulares rápidas frente a cambios ambientales. Cuando una célula recibe señales que requieren una producción inmediata de determinadas proteínas, puede incrementar la tasa de iniciación de la traducción sobre ARN mensajeros ya existentes. Esta estrategia proporciona una respuesta mucho más rápida que la síntesis de nuevos transcritos, permitiendo adaptaciones eficientes frente a estímulos fisiológicos o condiciones de estrés.
Otro aspecto fundamental es que los ribosomas no poseen especificidad inherente para tipos particulares de proteínas. Todos los ribosomas funcionales de una célula comparten esencialmente la misma maquinaria catalítica y pueden traducir cualquier ARN mensajero que contenga las señales apropiadas de iniciación y elongación. La especificidad de la proteína sintetizada está determinada exclusivamente por la secuencia de nucleótidos del ARN mensajero y no por características particulares del ribosoma. En consecuencia, el ribosoma actúa como una plataforma molecular universal capaz de ejecutar las reacciones químicas necesarias para formar enlaces peptídicos independientemente de la naturaleza de la proteína producida.
La función central del ribosoma consiste en catalizar la formación de enlaces peptídicos entre aminoácidos consecutivos. Esta actividad depende principalmente del ARN ribosómico presente en la subunidad mayor, lo que convierte al ribosoma en un ejemplo clásico de ribozima. Durante décadas se asumió que las proteínas ribosómicas eran las responsables directas de la catálisis; sin embargo, investigaciones posteriores demostraron que el centro peptidil transferasa está constituido fundamentalmente por ARN ribosómico, proporcionando evidencia sólida del papel catalítico de los ácidos nucleicos en la evolución biológica.
Los polisomas también tienen una relevancia evolutiva considerable. Su existencia permite que organismos de todos los dominios de la vida produzcan grandes cantidades de proteínas esenciales utilizando recursos limitados. Este principio de eficiencia contribuyó probablemente al éxito evolutivo de los sistemas celulares modernos, al maximizar la relación entre inversión energética y producción proteica.
En organismos procariotas, los polisomas pueden formarse incluso antes de que la transcripción haya concluido. Debido a la ausencia de compartimentalización nuclear, los ribosomas pueden unirse al ARN mensajero mientras este todavía está siendo sintetizado por la ARN polimerasa. Este fenómeno, denominado acoplamiento transcripción-traducción, incrementa aún más la velocidad de expresión génica y representa una característica distintiva de las bacterias.
En organismos eucariotas, aunque la transcripción y la traducción están separadas espacialmente por la envoltura nuclear, los polisomas siguen siendo esenciales para alcanzar elevadas tasas de producción proteica. Células altamente secretoras, como las plasmáticas productoras de anticuerpos, las células pancreáticas productoras de enzimas digestivas o los hepatocitos sintetizadores de proteínas plasmáticas, contienen abundantes polisomas asociados al retículo endoplásmico rugoso, reflejando su intensa actividad biosintética.
Los polirribosomas representan una organización molecular altamente eficiente que permite la traducción simultánea de una única molécula de ARN mensajero por múltiples ribosomas. Esta disposición incrementa la velocidad de síntesis proteica, optimiza el uso de recursos celulares, facilita la regulación de la expresión génica y garantiza una producción masiva de proteínas indispensable para el funcionamiento, crecimiento, diferenciación y supervivencia de las células.


Fuente y lecturas recomendadas:
- Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2022). Molecular Biology of the Cell (7th ed.). W. W. Norton & Company.
- Amunts, A., Brown, A., Bai, X. C., Llácer, J. L., Hussain, T., Emsley, P., Long, F., Murshudov, G., Scheres, S. H. W., & Ramakrishnan, V. (2015). Structure of the yeast mitochondrial large ribosomal subunit. Science, 343(6178), 1485–1489.
- Frank, J. (2017). Molecular Machines in Biology: Workshop of the Cell. Cambridge University Press.
- Kozak, M. (1999). Initiation of translation in prokaryotes and eukaryotes. Gene, 234(2), 187–208.
- Lodish, H., Berk, A., Kaiser, C. A., Krieger, M., Bretscher, A., Ploegh, H., Amon, A., & Martin, K. C. (2021). Molecular Cell Biology (9th ed.). W. H. Freeman.
- Milo, R., & Phillips, R. (2015). Cell Biology by the Numbers. Garland Science.
- Nissen, P., Hansen, J., Ban, N., Moore, P. B., & Steitz, T. A. (2000). The structural basis of ribosome activity in peptide bond synthesis. Science, 289(5481), 920–930.
- Shirokikh, N. E., & Preiss, T. (2018). Translation initiation by cap-dependent ribosome recruitment: Recent insights and open questions. Wiley Interdisciplinary Reviews: RNA, 9(4), e1473.
- Steitz, T. A. (2008). A structural understanding of the dynamic ribosome machine. Nature Reviews Molecular Cell Biology, 9(3), 242–253.
- Watson, J. D., Baker, T. A., Bell, S. P., Gann, A., Levine, M., & Losick, R. (2014). Molecular Biology of the Gene (7th ed.). Pearson.
- Weingarten-Gabbay, S., & Segal, E. (2014). The grammar of translation initiation in eukaryotes. Genome Biology, 15(10), 1–10.
Aprende administración paso a paso

ADMINISTRACION DESDE CERO