
La meiosis es un proceso especializado de división celular propio de los organismos eucariotas sexuales mediante el cual una célula diploide origina células haploides genéticamente distintas, lo cual constituye la base biológica de la reproducción sexual y de la estabilidad del número cromosómico entre generaciones. En el análisis contemporáneo de la biología celular, este proceso ha sido descrito como una secuencia altamente regulada de eventos coordinados de recombinación, segregación cromosómica y control del ciclo celular, tal como se detalla en la revisión integral sobre meiosis en mamíferos publicada por Handel y Schimenti en Nature Reviews Genetics en 2010, donde se enfatiza que la meiosis no es únicamente una división reductiva, sino un mecanismo de reorganización genómica esencial para la evolución y la fertilidad.
La importancia biológica de la meiosis radica en dos funciones fundamentales. En primer lugar, reduce el número cromosómico de diploide a haploide, lo cual permite que la fecundación restablezca la dotación cromosómica característica de la especie. En segundo lugar, genera variabilidad genética mediante recombinación homóloga y segregación independiente de cromosomas, procesos ampliamente caracterizados en la síntesis de Zickler y Kleckner publicada en Annual Review of Genetics en 2015, donde se establece que la recombinación meiótica no es un evento accesorio sino un requisito estructural para la correcta segregación cromosómica.
Base celular y preparación de la meiosis
Antes del inicio de la meiosis, las células germinales diploides humanas poseen cuarenta y seis cromosomas organizados en veintitrés pares homólogos. Cada cromosoma ha sido previamente replicado durante la fase S del ciclo celular, generando dos cromátidas hermanas unidas por cohesinas, lo cual produce un contenido total de ácido desoxirribonucleico equivalente a noventa y dos moléculas de doble hélice por célula. Este estado preparatorio es esencial porque la separación de homólogos y cromátidas requiere estructuras duplicadas, como se describe en el modelo clásico de organización cromosómica de Alberts en Molecular Biology of the Cell.
El inicio de la meiosis implica la activación de programas moleculares específicos que distinguen la línea germinal de la mitosis somática. De acuerdo con estudios mecanísticos sobre recombinación meiótica iniciados por Keeney en Genes and Development en 2001, la meiosis se inicia con la formación de roturas programadas en el ácido desoxirribonucleico mediadas por la proteína Spo11, evento indispensable para el apareamiento de cromosomas homólogos.
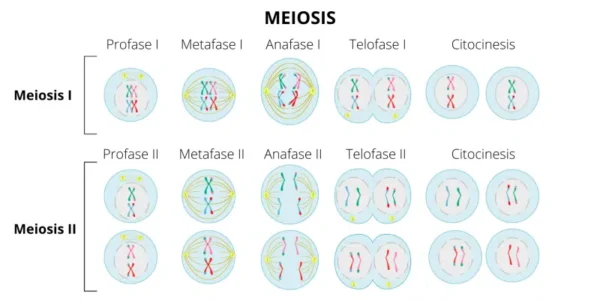
MEIOSIS I: división reduccional y reorganización genética
La meiosis I es una división reduccional porque separa cromosomas homólogos y reduce la ploidía de diploide a haploide, manteniendo cromátidas hermanas unidas. Este proceso ha sido descrito en detalle por Page y Hawley en Nature Reviews Genetics en 2003 como una etapa en la que la segregación cromosómica depende críticamente de la recombinación previa.
Profase I: organización compleja y recombinación genética
La profase I es la fase más extensa y compleja de la meiosis, caracterizada por la sinapsis de cromosomas homólogos, la formación del complejo sinaptonémico y la recombinación genética.
En leptoteno, los cromosomas comienzan su condensación progresiva y se hacen visibles como estructuras filamentosas individuales. Este proceso de condensación permite la búsqueda de homología entre cromosomas maternos y paternos, fenómeno descrito en estudios citológicos clásicos integrados en la síntesis moderna de Zickler y Kleckner en 2015.
Durante cigoteno, los cromosomas homólogos inician el apareamiento mediante sinapsis, proceso mediado por el ensamblaje del complejo sinaptonémico, una estructura proteica que estabiliza la alineación cromosómica. La formación de bivalentes en esta etapa es esencial para asegurar la recombinación precisa y prevenir errores de segregación.
En paquiteno ocurre la recombinación homóloga mediante entrecruzamiento entre cromátidas no hermanas. Este proceso es iniciado por roturas de doble cadena en el ácido desoxirribonucleico generadas por Spo11, como se describe en el modelo molecular de Keeney en 2001, y posteriormente reparadas mediante mecanismos de recombinación homóloga que producen intercambio genético estable, según lo detallado por Hunter en Annual Review of Genetics en 2015. Este intercambio es la base molecular del entrecruzamiento que genera nuevas combinaciones alélicas.
En diploteno comienza la separación progresiva de cromosomas homólogos, que permanecen unidos en regiones específicas denominadas quiasmas. Estos quiasmas representan los puntos físicos de recombinación y son esenciales para la estabilidad del bivalente, como se explica en la revisión de Page y Hawley en 2003, donde se destaca que la ausencia de quiasmas conduce a errores de segregación cromosómica.
En diacinesis los cromosomas alcanzan su máxima condensación, la envoltura nuclear se desorganiza y se forma el huso meiótico. Este estado prepara la célula para la segregación cromosómica ordenada, como parte del control estructural del ciclo meiótico descrito en Alberts Molecular Biology of the Cell.
Metafase I: alineación independiente y variabilidad genética
En metafase I, los bivalentes se alinean en el plano ecuatorial del huso meiótico. Cada cromosoma homólogo se orienta hacia polos opuestos de la célula, mientras las cromátidas hermanas permanecen unidas. La orientación aleatoria de los bivalentes constituye la base de la segregación independiente, un mecanismo fundamental de variabilidad genética descrito en la síntesis de Zickler y Kleckner en 2015.
Anafase I: separación de cromosomas homólogos
En anafase I ocurre la separación de cromosomas homólogos hacia polos opuestos sin ruptura de centrómeros. Este evento depende de la pérdida controlada de cohesión en brazos cromosómicos mientras se mantiene cohesión centromérica, un mecanismo regulado por complejos proteicos descritos por Petronczki y colaboradores en Cell en 2003. La segregación correcta en esta etapa es crítica para evitar aneuploidías.
Telofase I: reducción cromosómica
En telofase I los cromosomas alcanzan polos opuestos y puede reorganizarse parcialmente la envoltura nuclear. El resultado son dos células haploides con veintitrés cromosomas duplicados, cada uno constituido por dos cromátidas hermanas. Este estado intermedio es fundamental porque prepara la separación final de cromátidas en la segunda división meiótica.
MEIOSIS II: separación de cromátidas hermanas
La meiosis II es una división ecuacional similar a la mitosis en su mecanismo, pero ocurre en células haploides y sin replicación previa de ácido desoxirribonucleico. Este aspecto ha sido descrito en detalle en la revisión de Handel y Schimenti en 2010 como una etapa necesaria para la segregación final de cromátidas.
Profase II: reorganización del aparato mitótico
Durante profase II los cromosomas se condensan nuevamente y se forma el huso meiótico. Si existe envoltura nuclear residual, esta se desintegra. Este proceso prepara la segregación de cromátidas hermanas mediante la reactivación del sistema de microtúbulos.
Metafase II: alineación cromosómica individual
En metafase II los cromosomas se alinean individualmente en el plano ecuatorial. Cada cromátida hermana se conecta a microtúbulos provenientes de polos opuestos, garantizando su separación posterior.
Anafase II: separación de cromátidas hermanas
En anafase II ocurre la ruptura de cohesinas centroméricas, lo que permite la separación de cromátidas hermanas. Este proceso ha sido descrito por Petronczki en Cell en 2003 como un evento altamente regulado por proteólisis dirigida que asegura la segregación exacta del material genético.
Telofase II: formación de células haploides finales
En telofase II se reconstituye la envoltura nuclear y los cromosomas se descondensan. La citocinesis da lugar a cuatro células haploides genéticamente distintas, cada una con veintitrés cromosomas simples.

Resultado biológico y significado evolutivo
El resultado final de la meiosis es la formación de cuatro células haploides genéticamente únicas. Esta diversidad surge de tres mecanismos fundamentales: recombinación genética durante paquiteno, segregación independiente de cromosomas homólogos en metafase I y anafase I, y separación de cromátidas hermanas en meiosis II. Estos mecanismos han sido descritos como la base molecular de la variabilidad genética en organismos sexuales en la revisión de Hunter en 2015.
En la gametogénesis masculina, la meiosis ocurre de forma continua desde la pubertad, produciendo espermatozoides de manera sostenida. En la gametogénesis femenina, la meiosis presenta detenciones prolongadas: inicia durante el desarrollo embrionario, se detiene en diploteno hasta la madurez sexual, y la meiosis II solo se completa si ocurre fecundación. Este comportamiento diferencial ha sido descrito en detalle por Handel y Schimenti en 2010 como un mecanismo de regulación temporal del desarrollo reproductivo.

Fuente y lecturas recomendadas:
- Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2015). Molecular Biology of the Cell(6th ed.). Garland Science.
- Handel, M. A., & Schimenti, J. C. (2010). Genetics of mammalian meiosis: Regulation, dynamics and impact on fertility. Nature Reviews Genetics, 11(2), 124–136.
- Hunter, N. (2015). Meiotic recombination: The essence of heredity. Annual Review of Genetics, 49, 17–43.
- Keeney, S. (2001). Mechanism of meiotic recombination initiation by Spo11. Genes & Development, 15(3), 335–354.
- Page, S. L., & Hawley, R. S. (2003). Chromosome choreography: The meiotic ballet. Nature Reviews Genetics, 4(1), 23–33.
- Petronczki, M., Siomos, M. F., & Nasmyth, K. (2003). Un ménage à quatre: The molecular biology of chromosome segregation in meiosis. Cell, 112(4), 423–440.
- Zickler, D., & Kleckner, N. (2015). Recombination, pairing, and synapsis of homologs during meiosis. Annual Review of Genetics, 49, 239–275.
 Síguenos en X: @el_homomedicus y @enarm_intensivo Síguenos en instagram: homomedicus y en Treads.net como: Homomedicus
Síguenos en X: @el_homomedicus y @enarm_intensivo Síguenos en instagram: homomedicus y en Treads.net como: Homomedicus
Aprende administración paso a paso

ADMINISTRACION DESDE CERO

