Los hidratos de carbono constituyen uno de los principales sustratos energéticos de la célula y participan de manera decisiva en la organización estructural, la señalización molecular, la regulación metabólica y la supervivencia celular. Aunque las proteínas y los líidos representan la mayor parte de la masa seca celular, los hidratos de carbono mantienen una relevancia fisiológica desproporcionadamente elevada debido a su rápida disponibilidad metabólica y a su capacidad para integrarse en múltiples rutas bioquímicas esenciales. En la mayoría de las células humanas, la cantidad total de hidratos de carbono almacenados es relativamente pequeña y suele representar alrededor del 1 % de la masa celular total; sin embargo, en células musculares puede alcanzar aproximadamente el 3 %, y en hepatocitos hasta el 5-6 %, debido a la gran capacidad de estos tejidos para almacenar glucógeno.
La principal forma de hidrato de carbono utilizable por las células es la glucosa. Esta molécula circula continuamente disuelta en el líquido extracelular y en el plasma sanguíneo, lo que garantiza una disponibilidad inmediata para prácticamente todos los tejidos. La glucosa posee una importancia fisiológica extraordinaria porque muchas células dependen casi exclusivamente de ella para obtener energía. Las neuronas, por ejemplo, consumen grandes cantidades de glucosa debido a sus elevadas demandas energéticas y a la limitada capacidad cerebral para utilizar otros combustibles en condiciones fisiológicas normales. Asimismo, los eritrocitos dependen completamente de la glucólisis anaerobia porque carecen de mitocondrias.
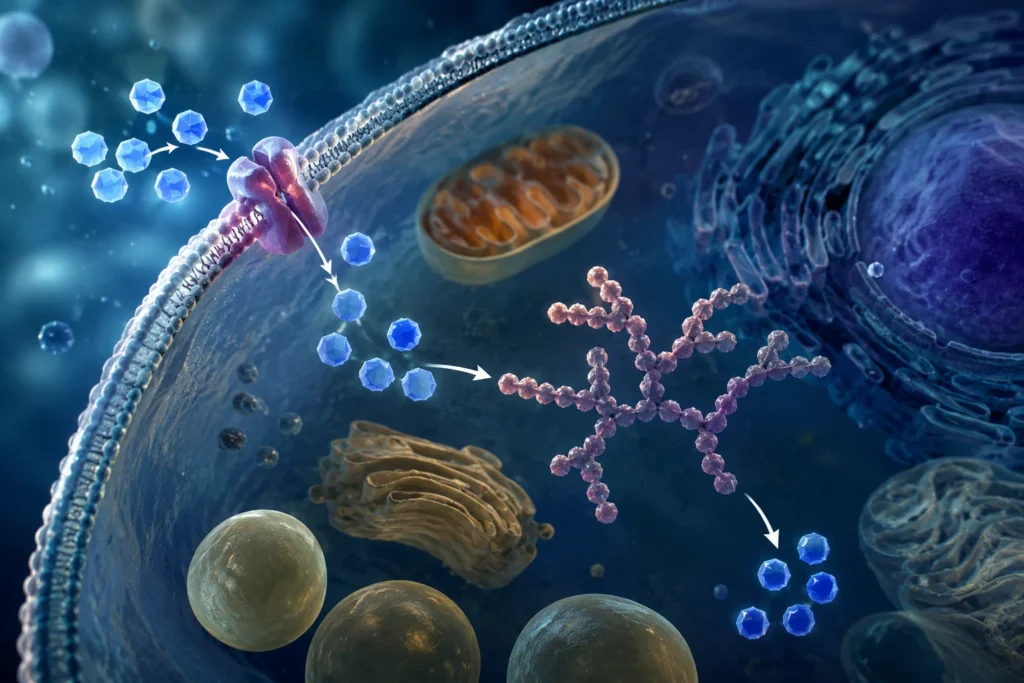
El ingreso de glucosa a la célula ocurre mediante proteínas transportadoras específicas denominadas GLUT. Una vez dentro del citoplasma, la glucosa es fosforilada a glucosa-6-fosfato por acción de la hexocinasa o de la glucocinasa, dependiendo del tejido. Esta fosforilación es esencial porque impide que la glucosa salga nuevamente de la célula y, además, la incorpora a múltiples vías metabólicas. La glucosa-6-fosfato puede ingresar en la glucólisis para producir ATP, puede desviarse hacia la vía de las pentosas fosfato para generar NADPH y ribosa-5-fosfato, o puede utilizarse para sintetizar glucógeno.
La importancia energética de los hidratos de carbono radica en que constituyen una fuente rápida y eficaz de ATP. Durante la glucólisis, una molécula de glucosa se transforma en 2 moléculas de piruvato con producción neta de ATP y NADH. Posteriormente, cuando existe oxígeno suficiente, el piruvato ingresa en la mitocondria y es oxidado mediante el ciclo de Krebs y la cadena respiratoria, procesos capaces de generar grandes cantidades de ATP. Esta producción energética resulta indispensable para mantener funciones celulares fundamentales como el transporte activo de membrana, la síntesis proteica, la contracción muscular, la transmisión nerviosa y los procesos de división celular.
Cuando la disponibilidad de glucosa excede las necesidades energéticas inmediatas, las células convierten parte de ella en glucógeno. El glucógeno es un polisacárido altamente ramificado formado por miles de moléculas de glucosa unidas mediante enlaces α-1,4 y α-1,6 glucosídicos. Su estructura extremadamente ramificada posee enorme relevancia funcional porque genera múltiples extremos terminales que permiten la liberación simultánea y rápida de glucosa durante períodos de elevada demanda energética.
La síntesis de glucógeno, denominada glucogenogénesis, ocurre principalmente en hígado y músculo esquelético. Este proceso requiere la participación coordinada de varias enzimas, incluyendo glucógeno sintasa y enzima ramificante. La degradación del glucógeno, conocida como glucogenólisis, es catalizada principalmente por la glucógeno fosforilasa y por enzimas desramificantes. La regulación hormonal de estos procesos es extraordinariamente precisa. La insulina favorece la síntesis de glucógeno después de las comidas, mientras que el glucagón y la adrenalina estimulan su degradación durante el ayuno o el ejercicio.
El glucógeno hepático y el glucógeno muscular poseen funciones fisiológicas diferentes. El hígado actúa como un reservorio sistémico de glucosa destinado al mantenimiento de la glucemia. Los hepatocitos contienen glucosa-6-fosfatasa, enzima que permite liberar glucosa libre hacia la circulación sanguínea. Gracias a ello, durante el ayuno el hígado puede mantener concentraciones adecuadas de glucosa plasmática para abastecer órganos altamente dependientes de este sustrato, especialmente el sistema nervioso central.
En contraste, el músculo esquelético utiliza su glucógeno exclusivamente para satisfacer sus propias necesidades energéticas. Las fibras musculares carecen prácticamente de glucosa-6-fosfatasa, por lo que no pueden exportar glucosa al torrente sanguíneo. Durante el ejercicio intenso, el glucógeno muscular es degradado rápidamente para generar ATP necesario para la contracción muscular. Esta capacidad explica por qué el agotamiento del glucógeno se asocia directamente con la fatiga muscular y con la disminución del rendimiento físico.
Aunque las reservas celulares de hidratos de carbono son relativamente pequeñas comparadas con las reservas lipídicas, su relevancia metabólica es enorme debido a su disponibilidad inmediata. Los lípidos proporcionan mayor cantidad total de energía, pero su movilización y utilización son más lentas y dependen intensamente del metabolismo oxidativo. En cambio, la glucosa puede metabolizarse rápidamente incluso en condiciones anaerobias, permitiendo una respuesta energética inmediata frente a incrementos súbitos de demanda metabólica.
Los hidratos de carbono también cumplen funciones estructurales fundamentales. Muchos componentes celulares contienen cadenas glucídicas unidas covalentemente a proteínas y lípidos, formando glucoproteínas y glucolípidos. Estas moléculas participan en la estabilidad de membranas, en la adhesión celular, en el reconocimiento inmunológico y en la comunicación intercelular. El glucocálix, capa rica en carbohidratos presente sobre la superficie celular, desempeña funciones esenciales en protección mecánica, señalización y reconocimiento molecular.
Además, diversos hidratos de carbono forman parte de moléculas biológicamente críticas. La ribosa y la desoxirribosa constituyen componentes esenciales de los ácidos nucleicos ARN y ADN. Sin estas pentosas sería imposible el almacenamiento y la transmisión de información genética. Asimismo, nucleótidos como ATP, GTP y NAD contienen azúcares en su estructura molecular, lo que demuestra que los hidratos de carbono participan directamente en prácticamente todos los procesos bioquímicos celulares.
La homeostasis de la glucosa constituye uno de los sistemas regulatorios más estrictos del organismo. Concentraciones excesivamente bajas de glucosa comprometen rápidamente la función neurológica y pueden provocar daño celular irreversible. Por otro lado, concentraciones elevadas y sostenidas favorecen glicación proteica, estrés oxidativo, inflamación y lesión vascular. Por ello, el metabolismo de los hidratos de carbono está regulado mediante complejos mecanismos hormonales y enzimáticos que coordinan absorción, almacenamiento, utilización y producción endógena de glucosa.
Las alteraciones en el metabolismo del glucógeno producen enfermedades conocidas como glucogenosis. Estas patologías derivan de defectos genéticos en enzimas responsables de la síntesis o degradación del glucógeno y generan acumulación anormal o incapacidad para movilizar reservas energéticas. Dependiendo de la enzima afectada, pueden aparecer hepatomegalia, hipoglucemia, debilidad muscular, intolerancia al ejercicio o cardiopatías. La existencia de estas enfermedades demuestra la importancia crítica que posee el metabolismo de los hidratos de carbono para la función celular y para la supervivencia del organismo.
Los hidratos de carbono representan mucho más que simples reservas energéticas. Constituyen elementos centrales de la fisiología celular porque proporcionan energía inmediata, participan en estructuras moleculares esenciales, regulan procesos de señalización y sostienen la homeostasis metabólica. La presencia continua de glucosa en el medio extracelular y la capacidad de almacenamiento en forma de glucógeno permiten que las células respondan de manera rápida y eficiente a variaciones en las demandas energéticas, garantizando así el funcionamiento integrado del organismo humano.


Fuente y lecturas recomendadas:
- Adeva-Andany, M. M., González-Lucán, M., Donapetry-García, C., Fernández-Fernández, C., & Ameneiros-Rodríguez, E. (2016). Glycogen metabolism in humans. BBA Clinical, 5, 85-100. https://doi.org/10.1016/j.bbacli.2016.02.001
- Berg, J. M., Tymoczko, J. L., Gatto, G. J., & Stryer, L. (2019). Biochemistry (9th ed.). W. H. Freeman.
- Ferrier, D. R. (2017). Lippincott Illustrated Reviews: Biochemistry (7th ed.). Wolters Kluwer.
- Murray, R. K., Bender, D. A., Botham, K. M., Kennelly, P. J., Rodwell, V. W., & Weil, P. A. (2018). Harper’s Illustrated Biochemistry (31st ed.). McGraw-Hill Education.
- Nielsen, J., Ørtenblad, N., Kristensen, J. M., Mohr, M., Rasmussen, P., Juel, C., & Wojtaszewski, J. F. P. (2018). The dynamic life of the glycogen granule. Journal of Biological Chemistry, 293(19), 7089-7098. https://doi.org/10.1074/jbc.R117.802843
- Olsson, K., & Saltin, B. (1970). Variation in total body stores of glycogen with diet and exercise. Acta Physiologica Scandinavica, 80(1), 11-18. https://doi.org/10.1111/j.1748-1716.1970.tb04764.x
- Roach, P. J., Depaoli-Roach, A. A., Hurley, T. D., & Tagliabracci, V. S. (2012). Glycogen and its metabolism: Some new developments and old themes. Biochemical Journal, 441(3), 763-787. https://doi.org/10.1042/BJ20111416
- Sun, R. C., & Denko, N. C. (2014). Hypoxic regulation of glycogen metabolism in cancer cells. Molecular Aspects of Medicine, 47-48, 1-14. https://doi.org/10.1016/j.mam.2015.11.006
- Vigh-Larsen, J. F., Ørtenblad, N., Spriet, L. L., Overgaard, K., & Mohr, M. (2021). Muscle glycogen metabolism and high-intensity exercise performance. Sports Medicine, 51(9), 1857-1874. https://doi.org/10.1007/s40279-021-01475-0
Aprende administración paso a paso

ADMINISTRACION DESDE CERO