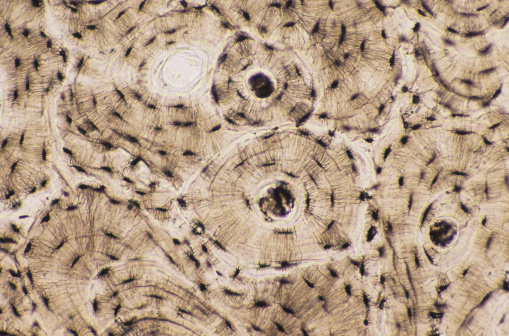
El tejido óseo se clasifica en hueso inmaduro (primario) y hueso maduro (secundario) según su grado de organización estructural.
Hueso maduro
El denominado hueso maduro, también conocido como hueso laminillar, presenta esta organización estructural específica no de manera arbitraria, sino como resultado de exigencias funcionales, mecánicas y metabólicas propias del tejido óseo altamente especializado.
La presencia de osteonas o sistemas de Havers responde a la necesidad de garantizar simultáneamente resistencia mecánica y viabilidad celular en un tejido intensamente mineralizado. Debido a que la matriz ósea posee una elevada concentración de sales de calcio y fosfato, lo que le confiere gran dureza, la difusión simple de nutrientes resulta insuficiente para mantener a las células. Por ello, el tejido se organiza en unidades cilíndricas alrededor de un conducto central que contiene vasos sanguíneos y fibras nerviosas. Esta disposición permite que cada osteona funcione como una unidad metabólica autónoma, en la que el aporte de oxígeno y nutrientes se distribuye de manera eficiente hacia los osteocitos.
En relación con lo anterior, los canalículos que irradian desde las lagunas donde se alojan los osteocitos no solo constituyen prolongaciones citoplasmáticas, sino que forman una red tridimensional de comunicación. Esta red es indispensable porque los osteocitos, al quedar atrapados en la matriz mineralizada, no pueden desplazarse. Por tanto, el patrón radial de los canalículos permite que las sustancias difundan desde el conducto central hacia todas las células, asegurando el intercambio metabólico y la homeostasis del tejido óseo.
Asimismo, la existencia de laminillas concéntricas con orientación alternante de las fibras colágenas tiene una justificación biomecánica fundamental. En cada laminilla, las fibras se disponen paralelas entre sí, pero cambian de dirección en las laminillas adyacentes. Esta disposición cruzada genera un efecto similar al de materiales compuestos laminados, lo que incrementa la resistencia frente a fuerzas de torsión, compresión y flexión. En consecuencia, el hueso no solo es rígido, sino también capaz de distribuir y disipar cargas mecánicas sin fracturarse fácilmente.
Por otra parte, las laminillas intersticiales representan vestigios de osteonas antiguas parcialmente reabsorbidas durante el proceso continuo de remodelación ósea. Esto evidencia que el hueso maduro no es una estructura estática, sino dinámica, en constante renovación. La presencia de estas laminillas refleja la adaptación del tejido a cambios mecánicos y metabólicos a lo largo del tiempo, optimizando su arquitectura interna.
En cuanto a los conductos de Volkmann, su disposición perpendicular o oblicua respecto al eje de las osteonas permite la interconexión entre distintos sistemas de Havers y la comunicación con las superficies interna y externa del hueso. Esta organización facilita una irrigación más amplia y eficiente, evitando zonas isquémicas dentro de un tejido tan denso. El hecho de que no estén rodeados por laminillas concéntricas se debe a que su función principal no es estructural, sino de conexión vascular.
Además, la orientación longitudinal de las osteonas paralela al eje del hueso largo no es casual. Esta alineación coincide con la dirección predominante de las fuerzas mecánicas que actúan sobre el hueso, especialmente durante la locomoción y el soporte del peso corporal. De esta manera, la arquitectura microscópica se adapta a las cargas macroscópicas, maximizando la resistencia con el menor gasto de material.
El hueso esponjoso maduro comparte la misma organización laminillar a nivel microscópico, pero se dispone en forma de trabéculas o espículas en lugar de estructuras compactas. Esta diferencia responde a una estrategia de optimización estructural: al crear espacios medulares intercomunicados, se reduce el peso del hueso sin comprometer su resistencia, y se facilita además el alojamiento de la médula ósea, esencial para la hematopoyesis.
Hueso inmaduro
El hueso inmaduro, también denominado hueso no laminillar o entretejido, presenta sus características particulares como consecuencia directa de su función primordial: formar tejido óseo de manera rápida durante etapas en las que la prioridad biológica no es la resistencia máxima, sino la generación expedita de una estructura inicial que posteriormente será perfeccionada.
La ausencia de una organización laminillar definida se explica por la velocidad con la que este tejido se sintetiza. Durante el desarrollo embrionario, así como en procesos de reparación, los osteoblastos producen matriz ósea de forma acelerada, lo que impide que las fibras colágenas se dispongan en patrones ordenados. En lugar de organizarse en capas paralelas, las fibras se depositan en múltiples direcciones, generando un entramado irregular. Esta disposición “entretejida” no responde a una deficiencia, sino a una estrategia biológica: construir rápidamente un armazón básico capaz de sostener el crecimiento o estabilizar una lesión.
La mayor cantidad de células por unidad de volumen en el hueso inmaduro se relaciona con su intensa actividad metabólica. Este tejido se encuentra en una fase de formación activa, por lo que contiene abundantes osteoblastos en proceso de síntesis de matriz y numerosos osteocitos recientemente incorporados. A diferencia del hueso maduro, donde la actividad celular es más equilibrada y regulada, el hueso inmaduro refleja un estado dinámico en el que predominan los procesos de producción sobre los de mantenimiento.
La distribución aleatoria de las células también deriva de esta formación acelerada. Dado que no existe todavía una arquitectura laminar establecida, los osteocitos quedan atrapados en la matriz en posiciones variables y sin una orientación preferente. En contraste, en el hueso maduro las células se alinean siguiendo la organización de las laminillas, lo que refleja una adaptación estructural más refinada. En el hueso inmaduro, en cambio, la disposición celular es un reflejo directo de un depósito matricial rápido y poco organizado.
Por otra parte, la mayor proporción de sustancia fundamental en la matriz del hueso inmaduro responde a un menor grado de mineralización. En las etapas iniciales, la matriz contiene una cantidad relativamente mayor de componentes orgánicos no mineralizados, como proteoglucanos y glicoproteínas. Esta composición favorece la flexibilidad y facilita la rápida expansión del tejido. Además, explica su afinidad tintorial por colorantes básicos, ya que la abundancia de componentes orgánicos y ácidos incrementa su basofilia, a diferencia del hueso maduro, cuya alta mineralización resalta la eosinofilia.
En relación con lo anterior, la mineralización incompleta del hueso inmaduro no constituye una limitación, sino una fase transitoria necesaria. La deposición de cristales minerales es un proceso progresivo que requiere tiempo. Inicialmente, una matriz menos mineralizada permite cierta elasticidad y adaptación a cambios morfológicos rápidos, lo cual es esencial durante el crecimiento fetal o la consolidación de fracturas. Con el tiempo, esta matriz se enriquece en minerales, aumentando su rigidez y transformándose en hueso maduro más resistente.
La rapidez de formación del hueso inmaduro es, por tanto, una de sus características más importantes y explica muchas de sus propiedades. Desde el punto de vista fisiológico, resulta más ventajoso generar primero un tejido estructuralmente básico en poco tiempo que esperar a la formación inmediata de un tejido altamente organizado. Posteriormente, mediante procesos de remodelación, este tejido inicial es reemplazado por hueso laminillar, que posee mejores propiedades mecánicas.
La presencia de hueso inmaduro en el adulto, especialmente en zonas de remodelación activa como los alvéolos dentarios o las inserciones tendinosas, se debe a que estos sitios están sometidos a microlesiones o a cambios mecánicos constantes. En tales condiciones, el organismo recurre nuevamente a la formación rápida de hueso entretejido como respuesta adaptativa inicial, que luego será reorganizado. Este fenómeno pone de manifiesto que el hueso inmaduro no es exclusivo del desarrollo embrionario, sino que constituye una fase universal en cualquier proceso de formación o reparación ósea.

Fuente y lecturas recomendadas:
- Ross, M. H. & Pawlina, W. (2020). Histología: texto y atlas: correlación con biología molecular y celular (8.ª ed.). Wolters Kluwer.
- Gartner, L. P. (2020). Textbook of Histology (5th ed.). Elsevier.
- Karp, G., Iwasa, J., & Marshall, W. (2019). Biología celular y molecular: conceptos y experimentos (8.ª ed.). McGraw-Hill Interamericana.
Aprende administración paso a paso

ADMINISTRACION DESDE CERO