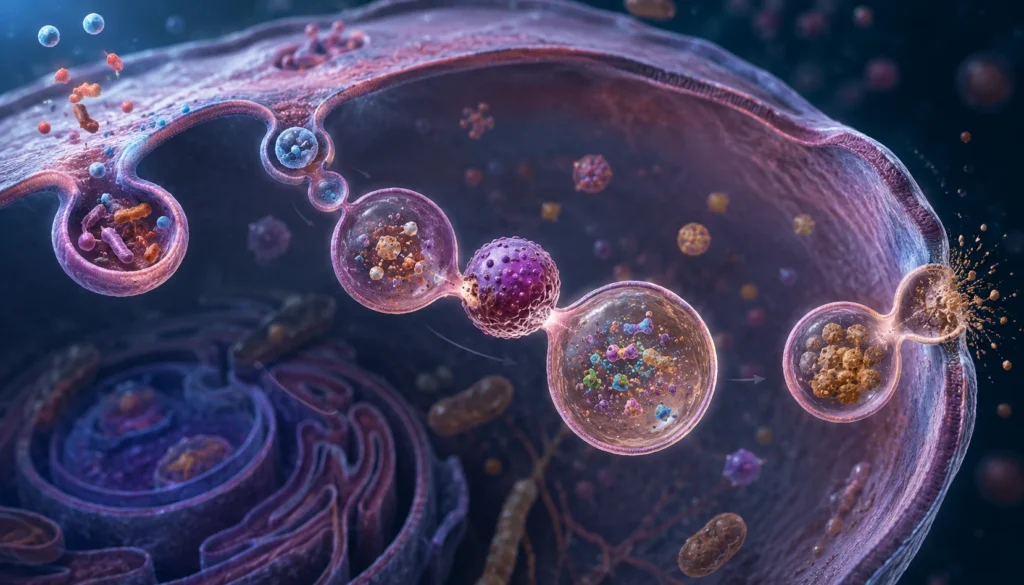
Los lisosomas constituyen los principales compartimentos degradativos de las células eucariotas y desempeñan una función esencial en la digestión intracelular de sustancias incorporadas desde el medio extracelular mediante pinocitosis y fagocitosis. Esta capacidad digestiva se fundamenta en la presencia de un amplio conjunto de enzimas hidrolíticas activas en un ambiente altamente ácido, así como en la capacidad de los lisosomas para fusionarse con vesículas endocíticas y fagocíticas que contienen material destinado a su degradación. La digestión lisosomal permite no solamente la eliminación de partículas extrañas, microorganismos y macromoléculas, sino también la recuperación de nutrientes y componentes reutilizables para el metabolismo celular.
La pinocitosis y la fagocitosis representan dos mecanismos especializados de endocitosis mediante los cuales la célula incorpora materiales desde el exterior. La pinocitosis permite la captación continua de líquidos extracelulares y de moléculas disueltas, mientras que la fagocitosis está especializada en la ingestión de partículas de gran tamaño, restos celulares, microorganismos y otros materiales sólidos. Como resultado de estos procesos se forman vesículas intracelulares delimitadas por membrana que contienen el material internalizado. Dichas vesículas constituyen compartimentos separados del citoplasma y requieren mecanismos adicionales para que su contenido pueda ser procesado y aprovechado por la célula.
Poco después de la formación de estas vesículas, ocurre una serie de eventos de tráfico intracelular que culminan con su interacción y posterior fusión con lisosomas. Este fenómeno depende de proteínas especializadas que regulan el reconocimiento, acercamiento y fusión de membranas biológicas. La unión de los lisosomas con vesículas endocíticas o fagocíticas genera estructuras híbridas denominadas endolisosomas o fagolisosomas, las cuales constituyen los verdaderos compartimentos donde ocurre la degradación activa de los materiales ingeridos. La formación de estos compartimentos garantiza que las enzimas lisosomales entren en contacto directo con los sustratos que deben ser degradados.
Los lisosomas contienen más de 60 tipos diferentes de hidrolasas ácidas capaces de degradar prácticamente todas las clases de macromoléculas biológicas. Entre estas enzimas se encuentran proteasas, lipasas, glucosidasas, nucleasas, fosfatasas y otras hidrolasas especializadas. Cada grupo enzimático actúa sobre un tipo específico de sustrato, permitiendo la degradación coordinada de proteínas, lípidos, polisacáridos, ácidos nucleicos y complejos macromoleculares. Gracias a esta extraordinaria diversidad enzimática, los lisosomas pueden procesar una enorme variedad de materiales procedentes tanto del exterior como del interior celular.
La actividad de estas enzimas depende críticamente de la existencia de un ambiente ácido en el interior lisosomal. El pH intraluminal se mantiene aproximadamente entre 4.5 y 5.0 mediante la acción de bombas de protones dependientes de ATP localizadas en la membrana lisosomal. Este ambiente ácido cumple dos funciones fundamentales. En primer lugar, proporciona las condiciones óptimas para la actividad catalítica de las hidrolasas ácidas. En segundo lugar, protege al citoplasma frente a una posible liberación accidental de enzimas lisosomales, ya que muchas de estas enzimas presentan una actividad significativamente menor a pH citoplasmático. La acidificación constituye, por tanto, un requisito indispensable para la digestión intracelular eficiente.
Una vez que el material captado entra en contacto con las hidrolasas lisosomales, se inicia un proceso progresivo de hidrólisis. Las proteínas son fragmentadas en péptidos y posteriormente en aminoácidos individuales. Los polisacáridos y otros hidratos de carbono complejos son convertidos en monosacáridos simples, incluyendo glucosa. Los lípidos son degradados hasta generar ácidos grasos, colesterol y otros componentes más pequeños. Los ácidos nucleicos son hidrolizados hasta producir nucleótidos, nucleósidos y bases nitrogenadas. Como consecuencia de estas reacciones, moléculas inicialmente demasiado grandes para ser utilizadas por la célula son transformadas en unidades moleculares simples susceptibles de ser reutilizadas metabólicamente.
La importancia fisiológica de esta degradación va mucho más allá de la simple eliminación de materiales extraños. Los productos generados por la digestión lisosomal constituyen una fuente importante de nutrientes celulares. Una vez obtenidos aminoácidos, monosacáridos, fosfatos, lípidos simples y otros metabolitos, estos son transportados a través de proteínas especializadas presentes en la membrana lisosomal hacia el citoplasma. Allí pueden incorporarse a rutas biosintéticas para la producción de nuevas macromoléculas o ingresar en vías catabólicas destinadas a la obtención de energía. En consecuencia, los lisosomas participan activamente en el reciclaje de componentes celulares y en el mantenimiento de la homeostasis metabólica.
El transporte de los productos de degradación desde la luz lisosomal hacia el citoplasma es un proceso altamente regulado. Diversos transportadores de membrana permiten la exportación selectiva de aminoácidos, azúcares, iones y otros metabolitos. Gracias a este sistema, la célula recupera recursos valiosos que de otro modo se perderían. Este mecanismo resulta particularmente importante en condiciones de escasez nutricional, durante las cuales la degradación lisosomal contribuye significativamente al suministro de nutrientes necesarios para la supervivencia celular.
Sin embargo, no todos los materiales internalizados pueden ser degradados completamente. Algunas sustancias poseen estructuras químicas extremadamente resistentes a la acción de las hidrolasas lisosomales. Entre ellas pueden encontrarse ciertos pigmentos, materiales minerales, compuestos sintéticos, partículas inorgánicas y residuos derivados de procesos metabólicos complejos. Los restos no degradados permanecen acumulados dentro de la vesícula digestiva formando una estructura denominada cuerpo residual. Este cuerpo residual representa el remanente final del proceso digestivo intracelular y contiene exclusivamente materiales que la célula no ha podido descomponer.
La acumulación de cuerpos residuales constituye una consecuencia natural de la digestión incompleta de determinados materiales. En muchas células, especialmente aquellas con una vida prolongada, los cuerpos residuales pueden acumularse progresivamente con el tiempo. Sin embargo, en la mayoría de los casos la célula dispone de mecanismos para eliminar estos residuos mediante exocitosis. Durante este proceso, la membrana de la vesícula residual se fusiona con la membrana plasmática y libera el contenido residual al espacio extracelular. Desde una perspectiva funcional, la exocitosis representa el proceso inverso a la endocitosis que originalmente permitió la entrada de las sustancias.
La exocitosis asociada a lisosomas y cuerpos residuales posee además una relevancia fisiológica considerable. No solamente permite eliminar materiales indigeribles, sino que también participa en procesos de remodelación tisular, reparación de membranas plasmáticas y mantenimiento de la integridad celular. La capacidad de los lisosomas para fusionarse con la membrana plasmática demuestra que estos orgánulos no son simplemente compartimentos degradativos, sino elementos dinámicos involucrados en múltiples aspectos de la fisiología celular.
Es así como las vesículas generadas por pinocitosis y fagocitosis, junto con los lisosomas que se fusionan con ellas, constituyen un auténtico sistema digestivo intracelular. De manera análoga a cómo el aparato digestivo de un organismo degrada alimentos para obtener nutrientes, el sistema endolisosomal degrada materiales incorporados por la célula para recuperar componentes útiles y eliminar residuos. Esta función resulta indispensable para la defensa frente a microorganismos, la eliminación de partículas extrañas, el reciclaje molecular, el mantenimiento del equilibrio metabólico y la supervivencia celular. La evidencia actual muestra además que los lisosomas actúan como centros de coordinación metabólica capaces de integrar la degradación de macromoléculas con mecanismos de señalización celular y regulación energética, lo que amplía considerablemente la visión clásica de estos orgánulos como simples compartimentos digestivos.


Fuente y lecturas recomendadas:
- Ballabio, A., & Settembre, C. (2024). Lysosomes as coordinators of cellular catabolism, metabolic signalling and organ physiology. Nature Reviews Molecular Cell Biology, 25(4), 223–245. https://doi.org/10.1038/s41580-023-00676-x
- Luzio, J. P., Pryor, P. R., & Bright, N. A. (2007). Lysosomes: Fusion and function. Nature Reviews Molecular Cell Biology, 8(8), 622–632. https://doi.org/10.1038/nrm2217
- Saftig, P., & Klumperman, J. (2009). Lysosome biogenesis and lysosomal membrane proteins: Trafficking meets function. Nature Reviews Molecular Cell Biology, 10(9), 623–635. https://doi.org/10.1038/nrm2745
- Settembre, C., Fraldi, A., Medina, D. L., & Ballabio, A. (2013). Signals from the lysosome: A control centre for cellular clearance and energy metabolism. Nature Reviews Molecular Cell Biology, 14(5), 283–296. https://doi.org/10.1038/nrm3565
- Tancini, B., Buratta, S., Delo, F., Sagini, K., Chiaradia, E., Pellegrino, R. M., Emiliani, C., & Urbanelli, L. (2020). Lysosomal exocytosis: The extracellular role of an intracellular organelle. Membranes, 10(12), 406. https://doi.org/10.3390/membranes10120406
- Xu, H., & Ren, D. (2015). Lysosomal physiology. Annual Review of Physiology, 77, 57–80. https://doi.org/10.1146/annurev-physiol-021014-071649
- Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2002). Lysosomes. En Molecular Biology of the Cell (4.ª ed.). Garland Science. NCBI Bookshelf. https://www.ncbi.nlm.nih.gov/books/NBK9953/
Aprende administración paso a paso

ADMINISTRACION DESDE CERO