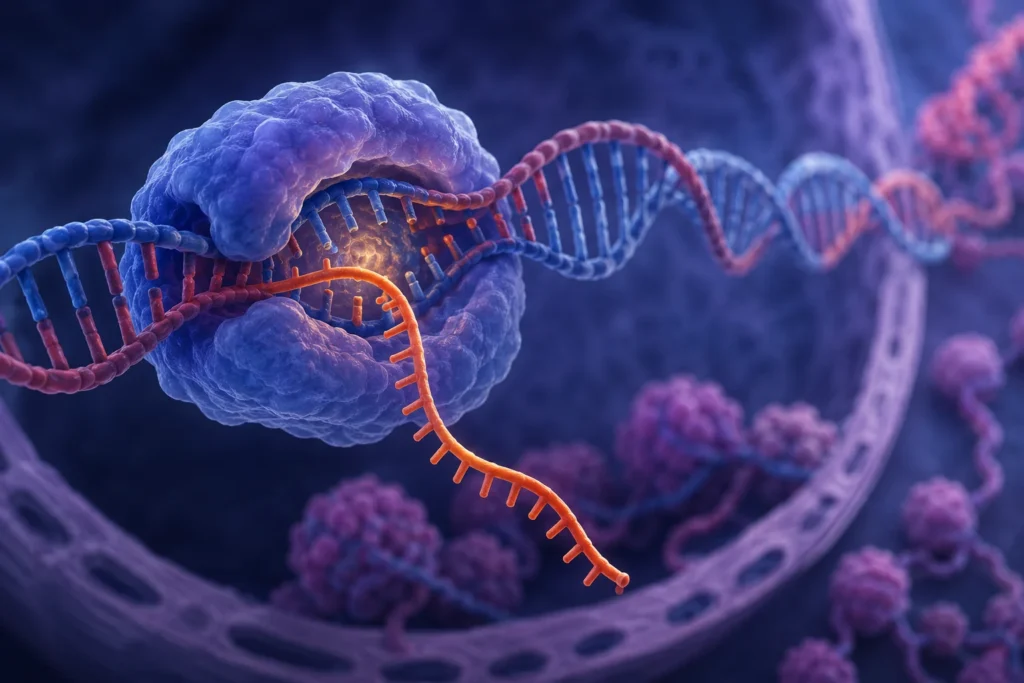
La síntesis de ARN a partir de una cadena de ADN, proceso denominado transcripción, constituye uno de los mecanismos fundamentales mediante los cuales la información genética almacenada en el ADN se convierte en moléculas funcionales capaces de dirigir la actividad celular. Este proceso permite transferir la información genética desde una secuencia de nucleótidos contenida en el ADN hacia una molécula de ARN complementaria, que posteriormente participará en la síntesis de proteínas o desempeñará funciones estructurales, reguladoras o catalíticas dentro de la célula. La transcripción ocurre principalmente en el núcleo de las células eucariotas y representa el primer paso de la expresión génica.
El ADN almacena la información genética en forma de secuencias específicas de nucleótidos organizadas en dos cadenas antiparalelas que forman una doble hélice. Para que esta información pueda utilizarse, resulta necesario copiar selectivamente determinados segmentos de ADN en moléculas de ARN. La utilización de una sola hebra como plantilla garantiza que la información genética se copie con precisión y que la secuencia resultante del ARN sea complementaria a la cadena molde y prácticamente idéntica a la cadena codificante, con la excepción de que el uracilo sustituye a la timina. Este mecanismo asegura la conservación de la información genética durante su transferencia desde el ADN hacia los productos funcionales de la célula.
Durante la transcripción, las dos hebras del ADN se separan de manera local y transitoria mediante la acción de la ARN polimerasa y de proteínas auxiliares. Esta separación genera una estructura denominada burbuja de transcripción, dentro de la cual queda expuesta la secuencia nucleotídica de la hebra molde. La ARN polimerasa reconoce secuencias específicas del ADN denominadas promotores, las cuales determinan el punto exacto donde debe comenzar la síntesis del ARN. Una vez unida al promotor, la enzima desenrolla una pequeña región de ADN y posiciona la hebra molde en su centro catalítico para iniciar la incorporación de nucleótidos complementarios.
La utilización de una sola hebra como plantilla posee una enorme importancia biológica. Si ambas hebras fueran copiadas simultáneamente para producir el mismo ARN, se generarían moléculas con secuencias diferentes y potencialmente incompatibles con la síntesis proteica normal. La selección específica de una hebra permite mantener una dirección definida de lectura genética y asegurar la producción correcta de moléculas funcionales. Además, diferentes genes pueden utilizar distintas hebras como molde dependiendo de su orientación dentro del cromosoma, lo que incrementa la capacidad de almacenamiento de información genética en el genoma.
La complementariedad entre bases nitrogenadas constituye el fundamento molecular de la síntesis de ARN. Cuando la ARN polimerasa avanza a lo largo de la hebra molde de ADN, incorpora nucleótidos de ARN siguiendo reglas específicas de apareamiento. La adenina presente en el ADN dirige la incorporación de uracilo en el ARN; la timina dirige la incorporación de adenina; la citosina dirige la incorporación de guanina; y la guanina dirige la incorporación de citosina. Estas interacciones se basan en la formación de puentes de hidrógeno específicos entre las bases nitrogenadas, lo que proporciona una elevada fidelidad durante la transcripción.
Los tripletes de nucleótidos presentes en la secuencia del ADN contienen la información necesaria para especificar la secuencia de aminoácidos de las proteínas. Durante la transcripción, dicha información es copiada en forma de codones dentro del ARN mensajero. Posteriormente, durante la traducción, los ribosomas interpretan estos codones y los convierten en secuencias de aminoácidos. De esta manera, la información genética fluye desde el ADN hacia el ARN y finalmente hacia las proteínas, siguiendo el principio fundamental de la biología molecular.
Los bloques básicos para la construcción del ARN son nucleótidos que presentan una organización química semejante a la de los nucleótidos del ADN. Cada nucleótido está compuesto por una base nitrogenada, un azúcar pentosa y uno o más grupos fosfato. Sin embargo, existen diferencias estructurales esenciales entre los nucleótidos de ADN y los de ARN que condicionan sus propiedades biológicas y químicas.
La primera diferencia corresponde al azúcar presente en cada molécula. Mientras que el ADN contiene desoxirribosa, el ARN contiene ribosa. La ribosa posee un grupo hidroxilo adicional unido al carbono 2’ de la pentosa. Esta pequeña modificación química tiene consecuencias funcionales profundas. La presencia del grupo hidroxilo aumenta la reactividad química del ARN, favorece la formación de estructuras tridimensionales complejas y permite que determinadas moléculas de ARN ejerzan funciones catalíticas. Sin embargo, también hace que el ARN sea más susceptible a la hidrólisis y, por tanto, menos estable que el ADN.
La menor estabilidad química del ARN resulta beneficiosa desde el punto de vista fisiológico. Muchas moléculas de ARN deben producirse rápidamente y degradarse cuando ya no son necesarias. Esta capacidad de renovación permite a las células responder con rapidez a cambios ambientales, señales hormonales, condiciones metabólicas y estímulos externos. En contraste, el ADN actúa como depósito permanente de información genética y requiere una estabilidad considerablemente mayor para preservar la integridad del genoma.
La segunda diferencia fundamental radica en la composición de las bases nitrogenadas. El ARN utiliza uracilo en lugar de timina. Ambas moléculas pertenecen al grupo de las pirimidinas y presentan estructuras químicas similares. Sin embargo, la timina posee un grupo metilo adicional que está ausente en el uracilo. La utilización de uracilo reduce el costo energético de la síntesis de ARN, ya que la producción metabólica de uracilo requiere menos modificaciones químicas. Dado que las moléculas de ARN suelen tener una vida media relativamente corta, la utilización de uracilo representa una estrategia eficiente desde el punto de vista evolutivo.
La sustitución de timina por uracilo también facilita los mecanismos celulares de reparación del ADN. La citosina puede sufrir desaminación espontánea y transformarse en uracilo. Si el ADN utilizara uracilo como base normal, sería difícil distinguir entre un uracilo legítimo y uno generado por daño químico. La presencia de timina en el ADN permite que los sistemas de reparación reconozcan cualquier uracilo presente como una anomalía que debe corregirse, contribuyendo así a la estabilidad genética.
La formación de nucleótidos de ARN sigue principios bioquímicos similares a los observados en la síntesis de nucleótidos de ADN. Los nucleótidos utilizados en la transcripción contienen las bases adenina, guanina, citosina y uracilo. Cada uno de ellos se encuentra unido a una molécula de ribosa y a grupos fosfato. Estas moléculas constituyen los sustratos directos empleados por la ARN polimerasa durante la elongación de la cadena de ARN.
Antes de incorporarse al ARN en crecimiento, los nucleótidos deben encontrarse en una forma activada denominada ribonucleósido trifosfato. Los principales sustratos son ATP, GTP, CTP y UTP. Cada uno contiene tres grupos fosfato unidos mediante enlaces fosfoanhidro de alta energía. Estos enlaces almacenan una cantidad significativa de energía química que será utilizada durante la formación de los enlaces fosfodiéster que conectan los nucleótidos consecutivos del ARN.
La activación de los nucleótidos es esencial porque la síntesis de ARN constituye un proceso termodinámicamente exigente. La ARN polimerasa aprovecha la energía liberada durante la ruptura de los enlaces entre los fosfatos terminales para impulsar la formación de nuevos enlaces covalentes dentro de la cadena naciente. Cuando un nucleótido es incorporado al ARN, se liberan pirofosfato y energía, lo que favorece que la reacción avance en dirección irreversible bajo condiciones fisiológicas.
La ARN polimerasa desempeña un papel central en todo el proceso. Esta enzima no solo cataliza la formación de enlaces fosfodiéster, sino que también reconoce promotores, desenrolla localmente el ADN, selecciona nucleótidos complementarios, corrige errores de incorporación y coordina múltiples factores reguladores. La extraordinaria precisión de la ARN polimerasa permite producir moléculas de ARN funcionales con una frecuencia relativamente baja de errores.
La síntesis del ARN ocurre exclusivamente en dirección 5’→3’. Esto significa que cada nuevo nucleótido se añade al extremo 3’ de la cadena en crecimiento. Simultáneamente, la ARN polimerasa se desplaza a lo largo de la hebra molde de ADN en dirección 3’→5’. Esta orientación antiparalela constituye una consecuencia directa de la estructura química de los ácidos nucleicos y de las características catalíticas de la enzima.
A medida que la ARN polimerasa avanza, la región de ADN situada detrás de la enzima vuelve a aparearse formando nuevamente la doble hélice original. De este modo, únicamente una pequeña porción del ADN permanece desenrollada en cada momento. Este mecanismo minimiza la exposición del material genético y contribuye a preservar su estabilidad estructural.
En las células eucariotas, la transcripción genera inicialmente un transcrito primario denominado pre-ARN mensajero. Esta molécula experimenta posteriormente diversas modificaciones antes de convertirse en ARN mensajero maduro. Entre estas modificaciones se incluyen la adición de una caperuza de 7-metilguanosina en el extremo 5’, la poliadenilación del extremo 3’ y la eliminación de intrones mediante empalme. Estos procesos incrementan la estabilidad del ARN, facilitan su transporte desde el núcleo hacia el citoplasma y permiten una regulación compleja de la expresión génica.
La síntesis de ARN representa, por tanto, un proceso extraordinariamente sofisticado que conecta la información genética almacenada en el ADN con la producción de proteínas y con numerosas funciones regulatorias esenciales para la vida celular. La participación coordinada de la ARN polimerasa, los nucleótidos activados, la complementariedad de bases y los mecanismos de procesamiento posteriores garantiza una transferencia precisa y eficiente de la información genética, permitiendo que cada célula exprese selectivamente los genes necesarios para mantener su estructura, metabolismo, diferenciación y capacidad de adaptación.


Fuente y lecturas recomendadas:
- Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2022). Molecular Biology of the Cell (7th ed.). W. W. Norton & Company.
- Cramer, P. (2019). Organization and regulation of gene transcription. Nature, 573(7772), 45–54. https://doi.org/10.1038/s41586-019-1517-4
- Fuchs, G., & Werner, F. (2021). RNA polymerases and the transcription machinery. Current Biology, 31(10), R487–R492. https://doi.org/10.1016/j.cub.2021.03.054
- Kornberg, R. D. (2007). The molecular basis of eukaryotic transcription. Proceedings of the National Academy of Sciences, 104(32), 12955–12961. https://doi.org/10.1073/pnas.0704138104
- Nelson, D. L., & Cox, M. M. (2021). Lehninger Principles of Biochemistry (8th ed.). W. H. Freeman.
- Watson, J. D., Baker, T. A., Bell, S. P., Gann, A., Levine, M., & Losick, R. (2014). Molecular Biology of the Gene (7th ed.). Pearson.
- Weinberg, D. E., & Grayhack, E. J. (2018). Post-transcriptional regulation of gene expression. Current Opinion in Genetics & Development, 48, 1–8. https://doi.org/10.1016/j.gde.2017.10.003
- Westover, K. D., Bushnell, D. A., & Kornberg, R. D. (2004). Structural basis of transcription: Nucleotide selection by rotation in the RNA polymerase II active center. Cell, 119(4), 481–489. https://doi.org/10.1016/j.cell.2004.10.016
- Yin, Y. W., & Steitz, T. A. (2004). The structural mechanism of translocation and helicase activity in T7 RNA polymerase. Cell, 116(3), 393–404. https://doi.org/10.1016/S0092-8674(04)00081-6
- Zhang, J., & Landick, R. (2016). A two-way street: Regulatory interplay between RNA polymerase and nascent RNA structure. Trends in Biochemical Sciences, 41(4), 293–310. https://doi.org/10.1016/j.tibs.2015.12.009
Aprende administración paso a paso

ADMINISTRACION DESDE CERO