La activación de los linfocitos T constituye uno de los mecanismos de control más estrictamente regulados de todo el sistema inmunitario adaptativo debido a que estas células poseen la capacidad de iniciar respuestas inflamatorias intensas, coordinar la actividad de numerosas poblaciones inmunológicas y destruir células propias infectadas o alteradas. Si el reconocimiento antigénico fuera el único requisito para su activación, el organismo desarrollaría respuestas inmunitarias permanentes contra proteínas propias, microorganismos comensales y una enorme cantidad de moléculas ambientales inocuas, lo que conduciría inevitablemente a enfermedades autoinmunes, inflamación crónica y daño tisular generalizado. Por esta razón, la evolución ha favorecido un sistema de activación basado en múltiples niveles de verificación, de manera que el reconocimiento del complejo formado por el antígeno y el complejo mayor de histocompatibilidad constituye solamente el primer requisito para la activación, pero no es suficiente por sí mismo para inducir una respuesta inmunitaria completa. La activación efectiva requiere la integración simultánea de señales antigénicas, señales coestimuladoras, citocinas y múltiples mecanismos reguladores que, en conjunto, permiten distinguir entre situaciones que representan un verdadero peligro biológico y aquellas que deben ser toleradas. Esta organización constituye uno de los principios fundamentales de la tolerancia inmunológica y de la discriminación entre lo dañino y lo inocuo.
La razón por la cual la presentación del complejo antígeno-complejo mayor de histocompatibilidad resulta insuficiente para activar completamente a un linfocito T radica en que las moléculas del complejo mayor de histocompatibilidad presentan de manera continua una enorme variedad de péptidos, independientemente de su origen. Estos péptidos pueden derivar de proteínas propias sintetizadas normalmente por el organismo, proteínas procedentes de microorganismos patógenos, proteínas provenientes de microorganismos comensales, proteínas alimentarias, alérgenos ambientales, restos celulares originados durante el recambio fisiológico de los tejidos e incluso proteínas modificadas por procesos metabólicos normales. En consecuencia, el simple hecho de que un péptido sea presentado sobre una molécula del complejo mayor de histocompatibilidad no proporciona información suficiente acerca de si dicho antígeno representa una amenaza para la integridad del organismo.
Este principio constituye una diferencia esencial entre el reconocimiento del antígeno y la interpretación inmunológica del mismo. El complejo mayor de histocompatibilidad funciona como un sistema de exhibición molecular que muestra fragmentos peptídicos a los linfocitos T, pero no transmite por sí mismo información acerca del contexto biológico en el cual se originó dicho péptido. La interpretación del contexto depende de señales adicionales generadas principalmente por las células presentadoras de antígeno profesionales después de detectar microorganismos o daño tisular.
Los autoantígenos representan un ejemplo particularmente ilustrativo de este fenómeno. Todas las células nucleadas sintetizan continuamente proteínas propias que son degradadas fisiológicamente por el proteasoma. Los fragmentos resultantes son cargados sobre moléculas del complejo mayor de histocompatibilidad clase I y presentados permanentemente en la superficie celular. Paralelamente, células dendríticas y macrófagos capturan proteínas liberadas durante el recambio normal de los tejidos y las presentan mediante moléculas del complejo mayor de histocompatibilidad clase II. Por lo tanto, los linfocitos T se encuentran expuestos continuamente a antígenos propios durante toda la vida.
Si el reconocimiento de estos complejos fuera suficiente para inducir activación, prácticamente todos los individuos desarrollarían respuestas inmunológicas permanentes contra proteínas estructurales, enzimas, receptores, moléculas del citoesqueleto y numerosos componentes celulares normales. La ausencia de estas respuestas refleja la existencia de mecanismos de tolerancia central y periférica que limitan la activación de linfocitos autorreactivos. Entre dichos mecanismos, la ausencia de señales coestimuladoras constituye uno de los más importantes.
Los antígenos ambientales benignos representan otra situación fisiológica que demuestra la necesidad de mecanismos adicionales de regulación. Cada día el organismo entra en contacto con miles de proteínas presentes en alimentos, polen, polvo, microorganismos comensales, microbiota intestinal, partículas inhaladas y otras moléculas ambientales incapaces de producir enfermedad. Muchas de estas proteínas son captadas por células dendríticas y posteriormente presentadas mediante moléculas del complejo mayor de histocompatibilidad. Sin embargo, en condiciones normales no inducen respuestas inmunitarias destructivas porque las células dendríticas permanecen en un estado funcional denominado tolerogénico, caracterizado por baja expresión de moléculas coestimuladoras y producción predominante de mediadores antiinflamatorios.
Este concepto dio origen al denominado modelo del peligro, según el cual el sistema inmunitario responde principalmente a señales asociadas con daño celular o infección y no simplemente a la presencia de moléculas extrañas. Cuando las células dendríticas detectan microorganismos mediante receptores especializados para reconocer patrones moleculares conservados o perciben moléculas liberadas por células lesionadas, experimentan un profundo proceso de maduración. Durante esta transición aumentan simultáneamente la expresión de moléculas del complejo mayor de histocompatibilidad, moléculas coestimuladoras y citocinas inflamatorias, adquiriendo capacidad para activar eficazmente linfocitos T vírgenes.
Entre las moléculas coestimuladoras más importantes destacan CD80 y CD86, también denominadas moléculas B7-1 y B7-2. Ambas pertenecen a la superfamilia de las inmunoglobulinas y se expresan principalmente sobre células dendríticas maduras, macrófagos activados y linfocitos B activados. En condiciones fisiológicas de ausencia de infección o inflamación, la expresión de estas moléculas permanece baja o prácticamente inexistente. Sin embargo, cuando las células presentadoras detectan microorganismos mediante receptores de reconocimiento de patrones o responden a citocinas inflamatorias, la síntesis de CD80 y CD86 aumenta de manera muy importante.
Este incremento constituye una señal biológica que informa al linfocito T que el antígeno está siendo presentado en un contexto potencialmente peligroso. En otras palabras, la presencia de CD80 y CD86 indica que la célula presentadora no solamente ha captado un antígeno, sino que además ha recibido evidencia molecular de infección, lesión tisular o inflamación suficiente para justificar el inicio de una respuesta inmunitaria adaptativa.
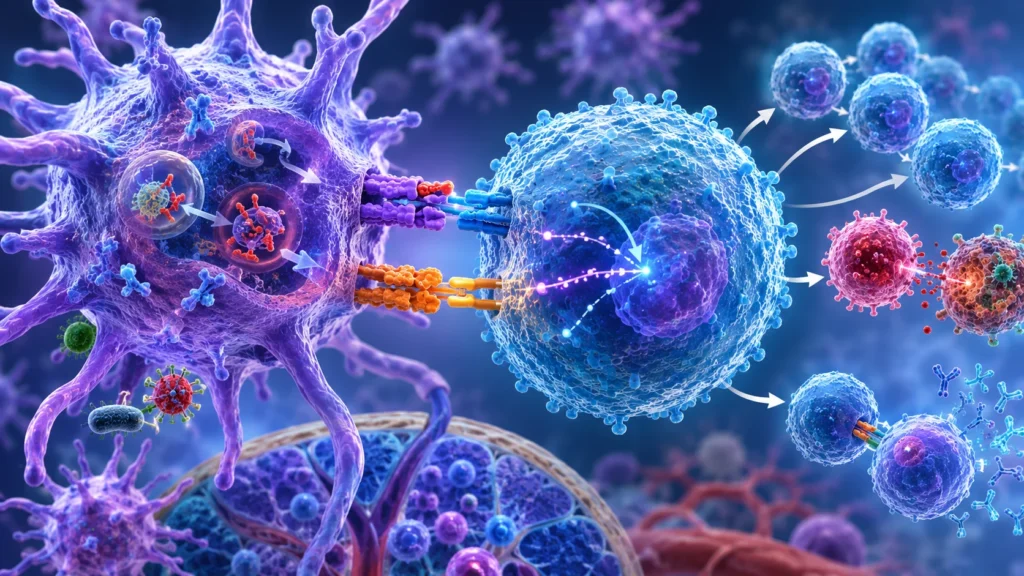
El principal receptor para estas moléculas en los linfocitos T vírgenes es CD28. Este receptor se expresa constitutivamente en la mayoría de los linfocitos T y constituye la molécula coestimuladora positiva más importante durante las fases iniciales de la respuesta inmunitaria. Cuando CD28 interactúa simultáneamente con CD80 o CD86 mientras el receptor del linfocito T reconoce el complejo antígeno-complejo mayor de histocompatibilidad, se desencadena una intensa amplificación de las vías de señalización intracelular.
Esta cooperación entre ambas señales resulta esencial porque el receptor del linfocito T genera principalmente señales relacionadas con la especificidad antigénica, mientras que CD28 aporta señales relacionadas con la intensidad, duración y viabilidad de la respuesta. Ninguna de las dos vías alcanza por sí sola la activación óptima del linfocito. La integración simultánea de ambas determina la activación coordinada de numerosos factores de transcripción responsables de iniciar el programa completo de diferenciación inmunológica.
La señal transmitida por CD28 reduce significativamente el umbral necesario para activar un linfocito T. Desde el punto de vista fisiológico, el umbral de activación representa la cantidad mínima de estímulo requerida para desencadenar una respuesta celular efectiva. Sin coestimulación, incluso el reconocimiento de un complejo antígeno-complejo mayor de histocompatibilidad de elevada afinidad suele resultar insuficiente para producir proliferación y diferenciación completas. En cambio, cuando CD28 recibe señales provenientes de CD80 o CD86, disminuye la intensidad de estímulo necesaria para activar múltiples vías intracelulares, facilitando que el linfocito responda rápida y eficazmente frente a microorganismos verdaderamente peligrosos.
La activación de CD28 potencia numerosas rutas de señalización intracelular, incluyendo las vías dependientes de fosfatidilinositol 3-cinasa, proteína cinasa B, proteína cinasa C, complejo diana mecanística de la rapamicina, proteína activadora 1, factor nuclear de células T activadas y factor nuclear κB. Estas vías inducen una profunda reorganización del metabolismo celular, incrementan la captación de glucosa, estimulan la síntesis de proteínas, favorecen la biogénesis mitocondrial y preparan al linfocito para experimentar múltiples divisiones celulares durante la expansión clonal.
Uno de los efectos biológicos más importantes de la señalización mediada por CD28 consiste en el aumento de la producción de interleucina 2. Esta citocina constituye el principal factor de crecimiento autocrino de los linfocitos T activados. Después de la activación inicial, el propio linfocito comienza a sintetizar grandes cantidades de interleucina 2 y simultáneamente incrementa la expresión de su receptor de alta afinidad. Como consecuencia, la interleucina 2 actúa sobre la misma célula que la produjo y sobre otras células vecinas del mismo clon, estimulando intensa proliferación, supervivencia y diferenciación.
La importancia de la interleucina 2 radica en que permite transformar el reconocimiento inicial de un número extremadamente pequeño de linfocitos específicos en una respuesta inmunitaria de gran magnitud. Antes de la exposición al antígeno puede existir únicamente un linfocito capaz de reconocer un determinado péptido. Gracias a la acción proliferativa de la interleucina 2, este único linfocito puede originar miles o millones de células hijas con idéntica especificidad, proporcionando suficiente capacidad efectora para controlar la infección.
Además de estimular la proliferación, la señalización mediada por CD28 incrementa la supervivencia celular mediante la inducción de proteínas antiapoptóticas pertenecientes a la familia Bcl-2, favorece la producción de otras citocinas inflamatorias, incrementa la expresión de moléculas de adhesión, estabiliza el ácido ribonucleico mensajero de diversas citocinas y promueve cambios metabólicos indispensables para sostener la intensa actividad biosintética de las células activadas.
Cuando el reconocimiento del antígeno ocurre en ausencia de coestimulación adecuada, el resultado suele ser completamente diferente. En lugar de proliferar y diferenciarse, el linfocito entra en un estado funcional denominado anergia. Durante este proceso la célula permanece viva, pero pierde la capacidad de responder eficazmente a exposiciones posteriores al mismo antígeno. La anergia constituye uno de los mecanismos más importantes de tolerancia periférica y contribuye significativamente a prevenir enfermedades autoinmunes.
En otros casos, la ausencia persistente de coestimulación puede inducir apoptosis o favorecer la diferenciación hacia linfocitos T reguladores, cuya función consiste precisamente en limitar respuestas inmunológicas excesivas y mantener la homeostasis inmunitaria. De esta manera, la falta de señales coestimuladoras no representa simplemente ausencia de activación, sino una instrucción biológica activa que orienta al sistema inmunitario hacia la tolerancia.
La importancia fisiológica de CD80, CD86 y CD28 queda demostrada por su relevancia clínica. Diversos tratamientos inmunomoduladores utilizados actualmente aprovechan este eje molecular para modificar la intensidad de la respuesta inmunitaria. La proteína de fusión abatacept bloquea la interacción entre CD80, CD86 y CD28, reduciendo la activación de linfocitos T en enfermedades autoinmunes como la artritis reumatoide. De manera inversa, numerosas estrategias de inmunoterapia contra el cáncer buscan potenciar la activación de linfocitos T favoreciendo la persistencia de señales coestimuladoras o bloqueando vías inhibitorias que contrarrestan la función de CD28.
La necesidad de señales coestimuladoras refleja uno de los principios más sofisticados de la inmunología moderna: el sistema inmunitario no responde únicamente al reconocimiento específico del antígeno, sino a la integración simultánea de información relacionada con la identidad del antígeno, el contexto inflamatorio, el estado funcional de la célula presentadora y las necesidades biológicas del organismo. La interacción entre CD80 y CD86 presentes sobre células dendríticas activadas y CD28 expresado en los linfocitos T constituye el mecanismo coestimulador más importante para reducir el umbral de activación, incrementar la producción de interleucina 2, promover la expansión clonal y asegurar que la respuesta inmunitaria adaptativa se inicie exclusivamente cuando exista evidencia suficiente de una amenaza real. Gracias a este sistema de múltiples verificaciones, el organismo logra un equilibrio extraordinariamente preciso entre una respuesta defensiva eficaz frente a microorganismos y la preservación de la tolerancia frente a autoantígenos y antígenos ambientales inocuos.

Fuente y lecturas recomendadas:
- Abbas, A. K., Lichtman, A. H., & Pillai, S. (2024). Cellular and Molecular Immunology (11th ed.). Elsevier.
- Bluestone, J. A. (1995). New perspectives of CD28-B7-mediated T cell costimulation. Immunity, 2(6), 555–559. https://doi.org/10.1016/1074-7613(95)90000-4
- Chen, L., & Flies, D. B. (2013). Molecular mechanisms of T cell co-stimulation and co-inhibition. Nature Reviews Immunology, 13(4), 227–242. https://doi.org/10.1038/nri3405
- Croft, M. (2003). Co-stimulatory members of the TNF receptor family: Keys to effective T-cell immunity? Nature Reviews Immunology, 3(8), 609–620. https://doi.org/10.1038/nri1148
- Lenschow, D. J., Walunas, T. L., & Bluestone, J. A. (1996). CD28/B7 system of T cell costimulation. Annual Review of Immunology, 14, 233–258. https://doi.org/10.1146/annurev.immunol.14.1.233
- Murphy, K., & Weaver, C. (2022). Janeway’s Immunobiology (10th ed.). W. W. Norton & Company.
- Sharpe, A. H., & Pauken, K. E. (2018). The diverse functions of the PD1 inhibitory pathway. Nature Reviews Immunology, 18(3), 153–167. https://doi.org/10.1038/nri.2017.108
- Steinman, R. M. (2012). Decisions about dendritic cells: Past, present, and future. Annual Review of Immunology, 30, 1–22. https://doi.org/10.1146/annurev-immunol-100311-102839
Aprende administración paso a paso

ADMINISTRACION DESDE CERO