La membrana celular constituye una de las estructuras más importantes para la vida porque establece el límite físico que separa el medio intracelular del medio extracelular y, al mismo tiempo, permite una comunicación continua y altamente regulada entre ambos compartimientos. Lejos de ser una simple envoltura pasiva, la membrana plasmática es una estructura dinámica, organizada y funcional que participa en prácticamente todos los procesos fisiológicos celulares, incluyendo el transporte de nutrientes, la eliminación de productos de desecho, la transmisión de señales, el reconocimiento celular, la adhesión intercelular, la regulación del volumen celular y el mantenimiento de los gradientes electroquímicos indispensables para la supervivencia de la célula. Su organización molecular responde a principios fisicoquímicos precisos que permiten combinar estabilidad estructural con una extraordinaria capacidad funcional.
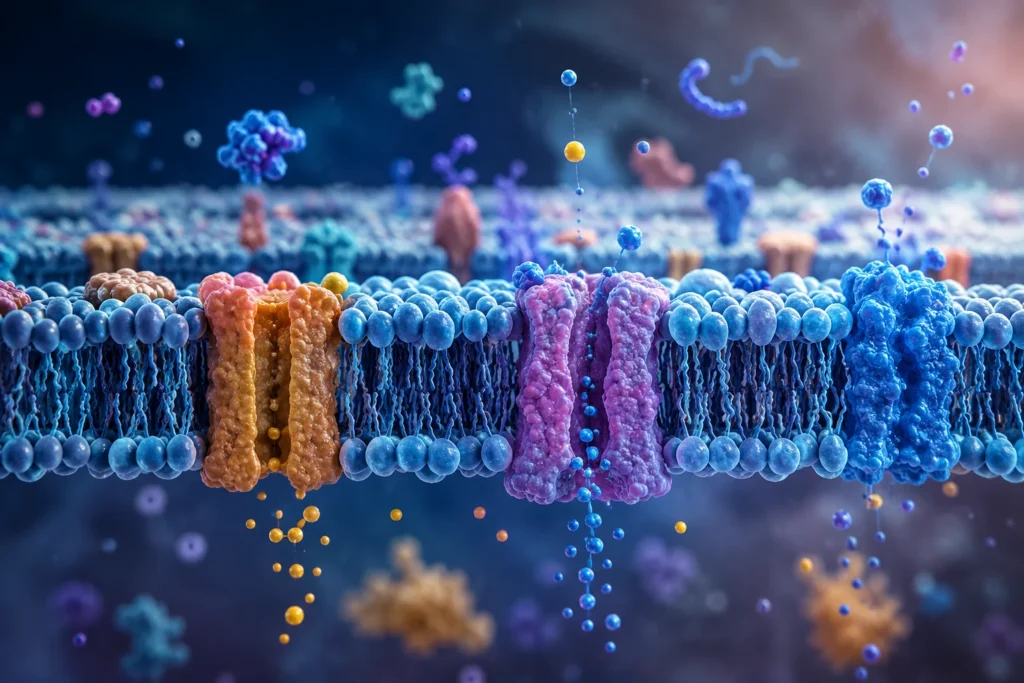
La arquitectura fundamental de la membrana celular está constituida por una bicapa lipídica en la que se encuentran incrustadas numerosas proteínas. Este concepto corresponde al modelo de mosaico fluido, según el cual los lípidos y las proteínas poseen movilidad lateral dentro del plano de la membrana, formando una estructura flexible y dinámica capaz de reorganizarse en respuesta a las necesidades fisiológicas de la célula. La bicapa está formada principalmente por fosfolípidos, colesterol y glucolípidos, mientras que las proteínas pueden encontrarse embebidas parcial o totalmente dentro de la membrana o asociadas a su superficie. Esta organización permite que la membrana funcione simultáneamente como barrera selectiva y como plataforma molecular para múltiples procesos celulares.
Los fosfolípidos representan el principal componente estructural de la bicapa. Cada molécula posee una región hidrofílica y una región hidrofóbica. La cabeza polar contiene grupos fosfato con afinidad por el agua, mientras que las colas hidrocarbonadas presentan características hidrofóbicas y tienden a evitar el contacto con el medio acuoso. Cuando estas moléculas se encuentran en un ambiente acuoso, se organizan espontáneamente en una bicapa debido a principios termodinámicos. Las cabezas hidrofílicas se orientan hacia los líquidos intra y extracelular, mientras que las colas hidrofóbicas se disponen hacia el interior de la membrana, formando un núcleo apolar que constituye la principal barrera para el paso de sustancias hidrosolubles.
Esta organización molecular explica por qué la bicapa lipídica es inmiscible tanto con el líquido intracelular como con el líquido extracelular. El agua es una molécula polar que forma extensas redes de enlaces de hidrógeno. Las cadenas hidrocarbonadas de los lípidos carecen de polaridad y no pueden establecer interacciones favorables con las moléculas de agua. Como consecuencia, la región central de la membrana actúa como una barrera energética que dificulta enormemente el movimiento espontáneo de sustancias polares o cargadas entre ambos compartimientos.
La impermeabilidad relativa de la bicapa lipídica es esencial para la vida celular. Si la membrana permitiera el libre intercambio de todas las sustancias, la composición química intracelular sería idéntica a la del medio extracelular, desapareciendo los gradientes de concentración y los potenciales eléctricos necesarios para la función celular. Gracias a la existencia de esta barrera, la célula puede mantener concentraciones intracelulares de potasio mucho mayores que las extracelulares, concentraciones extracelulares de sodio significativamente superiores a las intracelulares, niveles controlados de calcio citoplasmático y una composición metabólica propia. Estos gradientes constituyen la base de fenómenos fundamentales como la excitabilidad nerviosa, la contracción muscular, la secreción celular y el transporte secundario de nutrientes.
Las propiedades de permeabilidad de la bicapa dependen en gran medida de las características fisicoquímicas de las moléculas que intentan atravesarla. Las sustancias pequeñas y liposolubles presentan una elevada capacidad para difundirse directamente a través del núcleo hidrofóbico de la membrana. Entre ellas se encuentran el oxígeno, el dióxido de carbono, los gases anestésicos y numerosas hormonas esteroideas. Estas moléculas pueden disolverse transitoriamente en la fase lipídica y atravesar la membrana siguiendo sus gradientes de concentración sin necesidad de proteínas transportadoras.
Por el contrario, las moléculas hidrosolubles enfrentan una barrera considerable. La presencia de cargas eléctricas o de grupos polares genera interacciones desfavorables con el interior hidrofóbico de la bicapa, incrementando notablemente la energía necesaria para atravesarla. Los iones como sodio, potasio, calcio y cloruro prácticamente no pueden cruzar la membrana por difusión simple debido a sus cargas eléctricas y a las capas de hidratación que los rodean. De manera similar, moléculas polares como glucosa, aminoácidos, nucleótidos y numerosos metabolitos requieren mecanismos especializados para atravesar la membrana.
La existencia de proteínas insertadas en la bicapa lipídica resuelve este problema fisiológico fundamental. Las proteínas de membrana interrumpen la continuidad del núcleo hidrofóbico y proporcionan rutas específicas para el movimiento controlado de sustancias. Estas proteínas constituyen aproximadamente la mitad de la masa total de muchas membranas celulares y son responsables de la mayor parte de sus funciones fisiológicas.
Las proteínas integrales o transmembrana atraviesan parcial o completamente la bicapa lipídica. Sus regiones hidrofóbicas interactúan con los lípidos de la membrana, mientras que sus dominios hidrofílicos se proyectan hacia los medios intra y extracelular. Esta disposición les permite formar estructuras especializadas capaces de transportar moléculas, actuar como receptores, funcionar como enzimas o participar en mecanismos de adhesión celular.
Entre las proteínas transmembrana destacan las proteínas de canal. Estas proteínas forman poros acuosos que conectan ambos lados de la membrana. El interior del canal está revestido por residuos hidrofílicos que permiten el paso de agua o de determinadas especies iónicas sin necesidad de que estas entren en contacto con el núcleo hidrofóbico de la bicapa. De esta manera se reduce drásticamente la barrera energética que normalmente impediría el movimiento de sustancias polares.
Los canales iónicos constituyen algunos de los ejemplos más sofisticados de selectividad biológica. Aunque poseen diámetros extremadamente pequeños, son capaces de distinguir entre iones con características muy similares. Esta selectividad depende de regiones especializadas denominadas filtros de selectividad, donde la geometría molecular y la distribución de cargas permiten el paso de ciertos iones mientras excluyen otros. Algunos canales permiten principalmente el paso de potasio, otros de sodio, calcio o cloruro, contribuyendo así al mantenimiento de los gradientes electroquímicos celulares.
Además de la selectividad, muchos canales poseen mecanismos de compuerta que regulan su apertura y cierre. Algunos responden a cambios en el potencial eléctrico de membrana, otros a la unión de ligandos específicos y otros a estímulos mecánicos como el estiramiento celular. Esta capacidad reguladora convierte a los canales en elementos fundamentales para la excitabilidad neuronal, la conducción cardíaca, la contracción muscular, la secreción hormonal y numerosos procesos fisiológicos adicionales.
Un ejemplo particularmente importante de proteínas de canal corresponde a las acuaporinas. Estas proteínas permiten el paso extremadamente rápido de moléculas de agua a través de la membrana plasmática, facilitando el equilibrio osmótico entre los compartimientos intra y extracelular. La estructura de las acuaporinas permite el transporte eficiente de agua mientras impide el paso de protones y otros iones, preservando así los gradientes electroquímicos celulares.
Además de los canales, existen proteínas transportadoras o carriers. A diferencia de los canales, estas proteínas no generan un poro continuo entre ambos lados de la membrana. En lugar de ello, reconocen específicamente una molécula o ion determinado, se unen a él mediante sitios de reconocimiento molecular y experimentan cambios conformacionales que trasladan el sustrato desde un lado de la membrana hacia el otro.
El mecanismo de transporte mediado por proteínas transportadoras puede compararse con una puerta giratoria molecular. La proteína presenta inicialmente su sitio de unión hacia uno de los lados de la membrana. Tras unirse al sustrato, experimenta una reorganización estructural que expone dicho sitio hacia el lado opuesto, permitiendo la liberación de la molécula transportada. Posteriormente la proteína retorna a su conformación inicial para iniciar un nuevo ciclo de transporte.
La especificidad de las proteínas transportadoras es extraordinariamente elevada. Muchas reconocen únicamente una molécula determinada o un grupo muy restringido de compuestos estructuralmente relacionados. Esta especificidad surge de interacciones precisas entre el sustrato y los aminoácidos que forman el sitio de unión. La complementariedad geométrica y electroquímica asegura que únicamente determinadas moléculas puedan ser transportadas.
Las proteínas transportadoras participan tanto en mecanismos de transporte pasivo como en procesos de transporte activo. En el transporte facilitado, las moléculas se desplazan a favor de su gradiente electroquímico sin consumo directo de energía metabólica. Un ejemplo clásico es el transporte de glucosa mediado por la familia de transportadores GLUT, que permite el ingreso de glucosa a numerosas células del organismo.
En el transporte activo, las proteínas utilizan energía para mover sustancias contra sus gradientes de concentración. La fuente energética suele derivarse de la hidrólisis de ATP o del aprovechamiento de gradientes electroquímicos preexistentes. Un ejemplo fundamental es la ATPasa Na⁺/K⁺, que expulsa sodio hacia el exterior celular e introduce potasio al interior. Este mecanismo es esencial para mantener el potencial de membrana, el volumen celular y numerosos procesos de transporte secundario.
La presencia simultánea de una barrera lipídica y de proteínas altamente especializadas permite a la membrana celular ejercer un control extremadamente preciso sobre el intercambio de sustancias. La célula no depende únicamente de las propiedades físicas de la bicapa, sino también de una compleja red de proteínas reguladoras capaces de modificar continuamente la permeabilidad de la membrana según las necesidades fisiológicas.
La selectividad de canales y transportadores constituye una característica esencial para la homeostasis celular. Gracias a esta selectividad, la célula puede captar nutrientes específicos, eliminar metabolitos de desecho, regular sus concentraciones iónicas, responder a señales extracelulares y mantener condiciones internas compatibles con la vida. Sin estos mecanismos especializados, la organización bioquímica característica de las células desaparecería rápidamente.
Así la membrana celular representa una estructura altamente organizada en la que la bicapa lipídica proporciona una barrera física selectiva mientras que las proteínas de canal y las proteínas transportadoras generan rutas controladas para el movimiento de agua, iones y moléculas específicas. La interacción coordinada entre estos componentes permite mantener la homeostasis, la comunicación celular y el funcionamiento integrado de todos los tejidos del organismo.


Fuente y lecturas recomendadas:
- Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2022). Molecular biology of the cell (7th ed.). W. W. Norton & Company.
- Agre, P., King, L. S., Yasui, M., Guggino, W. B., Ottersen, O. P., Fujiyoshi, Y., Engel, A., & Nielsen, S. (2002). Aquaporin water channels—From atomic structure to clinical medicine. Journal of Physiology, 542(1), 3–16.
- Hille, B. (2001). Ion channels of excitable membranes (3rd ed.). Sinauer Associates.
- Lodish, H., Berk, A., Kaiser, C. A., Krieger, M., Bretscher, A., Ploegh, H., Amon, A., & Martin, K. C. (2021). Molecular cell biology (9th ed.). W. H. Freeman.
- Phillips, R., Kondev, J., Theriot, J., Garcia, H., & Orme, N. (2023). Physical biology of the cell(3rd ed.). Garland Science.
- Singer, S. J., & Nicolson, G. L. (1972). The fluid mosaic model of the structure of cell membranes. Science, 175(4023), 720–731.
- Uversky, V. N. (2019). Intrinsically disordered proteins and their “mysterious” membrane interactions. Frontiers in Physics, 7, 10.
- von Heijne, G. (2007). The membrane protein universe: What’s out there and why bother? Journal of Internal Medicine, 261(6), 543–557.
- Venable, R. M., Brown, F. L. H., & Pastor, R. W. (2015). Mechanical properties of lipid bilayers from molecular dynamics simulation. Chemical Society Reviews, 44(14), 4899–4914.
- Wilkens, S. (2015). Structure and mechanism of ABC transporters. F1000Prime Reports, 7, 14.
Aprende administración paso a paso

ADMINISTRACION DESDE CERO