La supervivencia celular depende de que el líquido extracelular conserve concentraciones relativamente estables de oxígeno y dióxido de carbono, porque ambos gases participan de manera directa en el metabolismo energético, en el equilibrio ácido-base y en la continuidad de múltiples procesos bioquímicos esenciales. Aunque el oxígeno y el dióxido de carbono son moléculas pequeñas y difunden con facilidad a través de las membranas biológicas, su regulación fisiológica no ocurre por simple difusión pasiva, sino mediante sistemas integrados de transporte, amortiguación, detección y respuesta que involucran la función pulmonar, la circulación sanguínea, la hemoglobina, el control neural de la ventilación y la regulación renal. La importancia de esta regulación es tan crítica que desviaciones relativamente pequeñas en las presiones parciales de estos gases pueden comprometer la producción de energía celular, alterar la función enzimática y poner en riesgo la viabilidad del organismo entero.
El oxígeno es el aceptor final de electrones en la cadena respiratoria mitocondrial. Esto significa que prácticamente toda la producción aerobia de adenosín trifosfato depende de su disponibilidad adecuada. Si la concentración de oxígeno en el líquido extracelular disminuye de manera importante, la fosforilación oxidativa se compromete, la célula reduce su producción energética y comienza a depender de vías anaerobias menos eficientes, con acumulación de lactato y alteración del metabolismo celular. Por esta razón, el organismo posee mecanismos extremadamente refinados para mantener una disponibilidad de oxígeno suficientemente constante en los tejidos, a pesar de las variaciones en el consumo metabólico, en la actividad física, en la altitud o en los cambios hemodinámicos.
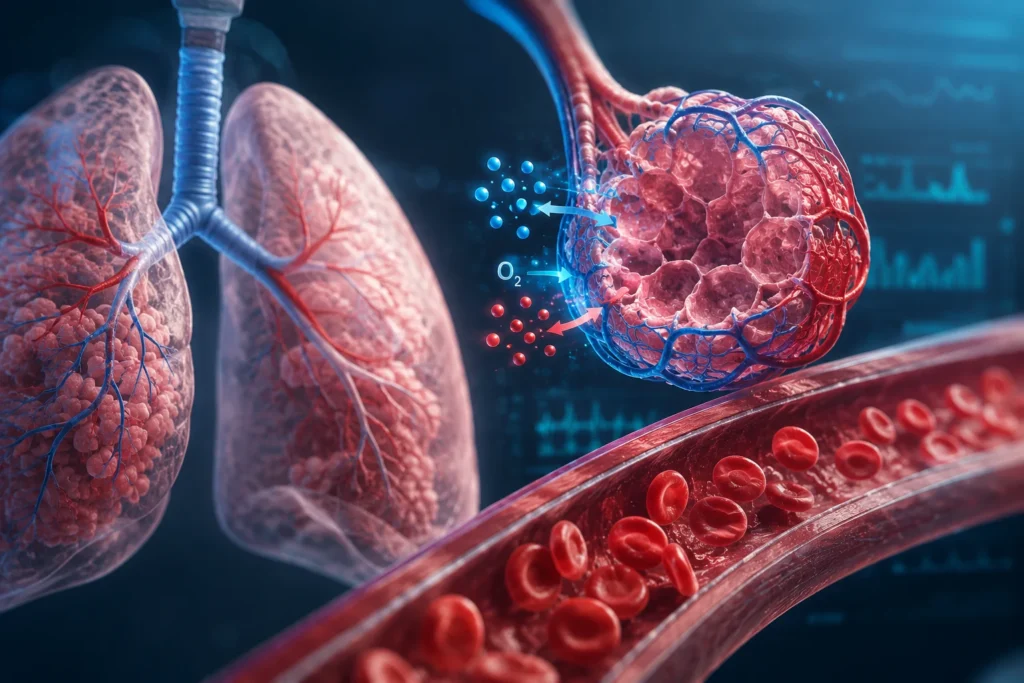
La regulación de la concentración de oxígeno en el líquido extracelular se basa, en primer lugar, en la captación pulmonar. Cuando el aire inspirado llega a los alvéolos, el oxígeno difunde a través de la membrana alveolocapilar siguiendo su gradiente de presión parcial hacia la sangre venosa que llega a los capilares pulmonares. Sin embargo, la cantidad de oxígeno que puede disolverse directamente en el plasma es muy pequeña, debido a la limitada solubilidad del gas en el agua. Si el transporte dependiera solo del oxígeno disuelto, la sangre sería incapaz de aportar suficiente oxígeno a los tejidos metabólicamente activos. Por ello, la evolución fisiológica desarrolló un sistema especializado de transporte basado en la hemoglobina contenida en los eritrocitos, que multiplica de manera extraordinaria la capacidad sanguínea de transporte de oxígeno.
La hemoglobina posee cuatro grupos hemo capaces de unirse reversiblemente al oxígeno. Esta unión no es rígida ni lineal, sino cooperativa. Cuando una molécula de oxígeno se une a un grupo hemo, aumenta la afinidad de los otros sitios por el oxígeno. Esta propiedad genera la característica curva sigmoidea de disociación oxígeno-hemoglobina. La trascendencia fisiológica de esta curva es enorme, porque permite que la hemoglobina se cargue de manera muy eficiente en los pulmones, donde la presión parcial de oxígeno es alta, y que descargue oxígeno en los tejidos, donde la presión parcial disminuye. De este modo, la hemoglobina no solo transporta oxígeno: actúa como un verdadero sistema amortiguador fisiológico de la concentración de oxígeno en el líquido extracelular.
Se habla de función amortiguadora de oxígeno porque la hemoglobina evita grandes fluctuaciones de oxígeno disuelto en los tejidos. Si la presión parcial de oxígeno en el líquido intersticial es adecuada, la hemoglobina conserva una fracción importante de su oxígeno unido y libera relativamente poco. En cambio, cuando la presión parcial disminuye por aumento del consumo metabólico, la afinidad funcional de la hemoglobina favorece una liberación mayor. Esto significa que el sistema responde automáticamente a la necesidad tisular. No se trata simplemente de un transporte pasivo, sino de un mecanismo químico de regulación dinámica que mantiene una disponibilidad relativamente estable de oxígeno en el entorno extracelular, incluso cuando la demanda cambia.
La capacidad reguladora de la hemoglobina se modifica además por varios factores fisiológicos. El aumento de dióxido de carbono, el incremento de protones, la elevación de la temperatura y el aumento de 2,3-bisfosfoglicerato desplazan la curva de disociación hacia la derecha, reduciendo la afinidad de la hemoglobina por el oxígeno y facilitando su liberación en tejidos metabólicamente activos. Este fenómeno, conocido como efecto Bohr, constituye un mecanismo de ajuste fino que conecta el metabolismo tisular con la entrega local de oxígeno. Un tejido que produce más dióxido de carbono y protones, precisamente porque está trabajando más intensamente, genera las condiciones químicas necesarias para recibir mayor cantidad de oxígeno. Así, la regulación del oxígeno no depende solo de la presión parcial, sino también del microambiente metabólico local.
Otro componente fundamental de la regulación del oxígeno en el líquido extracelular es el control ventilatorio mediado por quimiorreceptores periféricos, especialmente los cuerpos carotídeos. Cuando la presión parcial arterial de oxígeno cae de manera importante, estas estructuras detectan la disminución y aumentan la ventilación pulmonar. El incremento ventilatorio eleva la entrada de oxígeno alveolar y favorece la restauración de la oxigenación sanguínea. Sin embargo, en condiciones fisiológicas normales, el control ventilatorio está mucho más influido por el dióxido de carbono que por el oxígeno, lo cual refleja la importancia crítica del control del equilibrio ácido-base.
La regulación del dióxido de carbono en el líquido extracelular ocurre por mecanismos muy diferentes. Mientras que el oxígeno se regula principalmente mediante transporte amortiguado por hemoglobina y ajustes ventilatorios secundarios, el dióxido de carbono se controla mediante un sistema de retroalimentación ventilatoria extraordinariamente sensible. Esto se debe a que el dióxido de carbono no es solo un producto metabólico final; también es un determinante central del pH corporal a través de su conversión en ácido carbónico y protones. Una acumulación de dióxido de carbono no solo alteraría el intercambio gaseoso, sino que provocaría acidosis respiratoria y afectaría de forma profunda la función enzimática, neurológica y cardiovascular.
Las células producen dióxido de carbono de manera continua durante la oxidación de sustratos energéticos en la mitocondria. Este gas difunde desde el interior celular hacia el líquido intersticial, luego a la sangre capilar y finalmente hacia los pulmones. En la sangre, el dióxido de carbono se transporta de tres maneras: una pequeña fracción disuelta en plasma, una fracción unida a proteínas y hemoglobina en forma de compuestos carbamino, y la mayor parte convertida en bicarbonato mediante la acción de la anhidrasa carbónica dentro de los eritrocitos. Este último mecanismo es crucial porque permite transportar grandes cantidades de dióxido de carbono sin elevar excesivamente su presión parcial libre. Además, la hemoglobina desoxigenada puede aceptar protones, contribuyendo a amortiguar las alteraciones del pH durante este proceso.
La regulación fisiológica del dióxido de carbono depende principalmente de la detección de cambios en su presión parcial arterial por los quimiorreceptores centrales localizados en el tronco encefálico y por quimiorreceptores periféricos en cuerpos carotídeos y aórticos. El dióxido de carbono difunde con facilidad al líquido cefalorraquídeo, donde se hidrata y genera protones. El aumento local de protones estimula neuronas quimiosensibles que activan el centro respiratorio, aumentando tanto la frecuencia como la profundidad respiratoria. Este incremento de la ventilación alveolar favorece una mayor eliminación de dióxido de carbono y reduce su concentración hasta restablecer el equilibrio. Se trata de un clásico sistema de retroalimentación negativa de altísima sensibilidad.
La razón por la cual el dióxido de carbono tiene una influencia tan potente sobre la ventilación es que pequeños aumentos en su presión parcial generan cambios significativos en la concentración de protones, alterando el pH del líquido extracelular. El sistema respiratorio responde rápidamente porque la homeostasis ácido-base es incompatible con grandes oscilaciones. En condiciones normales, un ligero incremento de dióxido de carbono aumenta de forma casi inmediata la ventilación, y una disminución produce el efecto opuesto. De esta manera, la ventilación alveolar actúa como el principal mecanismo de regulación aguda de la concentración de dióxido de carbono en el líquido extracelular.
Existe además una relación íntima entre oxígeno y dióxido de carbono en la hemoglobina. Cuando la hemoglobina libera oxígeno en los tejidos, aumenta su capacidad para transportar dióxido de carbono y protones, fenómeno conocido como efecto Haldane. En los pulmones ocurre el proceso inverso: la unión de oxígeno a la hemoglobina reduce su afinidad por dióxido de carbono y protones, facilitando la eliminación pulmonar del gas. Este acoplamiento bioquímico mejora simultáneamente la entrega de oxígeno y la eliminación de dióxido de carbono, integrando ambos procesos en una regulación altamente eficiente.
En términos fisiopatológicos, el fracaso en la regulación de cualquiera de estos gases tiene consecuencias profundas. La hipoxemia sostenida compromete la respiración mitocondrial, induce disfunción celular y puede llevar a lesión tisular irreversible. La hipercapnia produce acidosis respiratoria, depresión neurológica y alteraciones hemodinámicas. La hipocapnia excesiva, por su parte, desplaza el equilibrio ácido-base hacia alcalosis, modifica la perfusión cerebral y altera la liberación de oxígeno desde la hemoglobina. Esto demuestra que la homeostasis gaseosa no consiste simplemente en aportar oxígeno o eliminar dióxido de carbono, sino en mantener una relación cuantitativa precisa entre transporte, metabolismo, ventilación y amortiguación química.
La regulación de las concentraciones de oxígeno y dióxido de carbono en el líquido extracelular constituye un sistema integrado de control homeostático en el que el oxígeno se mantiene principalmente mediante transporte amortiguado por hemoglobina y ajustes ventilatorios cuando la disponibilidad disminuye, mientras que el dióxido de carbono se regula sobre todo mediante un control ventilatorio rápido y sensible guiado por quimiorreceptores y estrechamente vinculado al equilibrio ácido-base. La hemoglobina no es solamente una molécula transportadora, sino un auténtico sistema químico regulador que conecta el intercambio pulmonar, el metabolismo tisular y la homeostasis global del organismo.


Fuente y lecturas recomendadas:
- West, J. B., & Luks, A. M. (2021). West’s respiratory physiology: The essentials (11th ed.). Wolters Kluwer.
- Hall, J. E. (2021). Guyton and Hall textbook of medical physiology (14th ed.). Elsevier.
- Levitzky, M. G. (2018). Pulmonary physiology (9th ed.). McGraw-Hill Education.
- Wagner, P. D. (2015). The physiological basis of pulmonary gas exchange: Implications for clinical interpretation of arterial blood gases. European Respiratory Journal, 45(1), 227–243.
- Weissmann, N., Sommer, N., Voswinckel, R., & Seeger, W. (2006). Oxygen sensors in hypoxic pulmonary vasoconstriction. Cardiovascular Research, 71(4), 620–629.
- Maren, T. H. (1967). Carbonic anhydrase: Chemistry, physiology, and inhibition. Physiological Reviews, 47(4), 595–781.
- Forster, R. E. (1964). Effect of dilution on blood gases and pH. Handbook of Physiology, Section 3: Respiration, 2, 801–816.
- Nattie, E., & Li, A. (2012). Central chemoreceptors: Locations and functions. Comprehensive Physiology, 2(1), 221–254.
- Winslow, R. M. (2007). The role of hemoglobin in oxygen transport. Microcirculation, 14(8), 689–700.
- Mairbäurl, H., & Weber, R. E. (2012). Oxygen transport by hemoglobin. Comprehensive Physiology, 2(2), 1463–1489.
Aprende administración paso a paso

ADMINISTRACION DESDE CERO