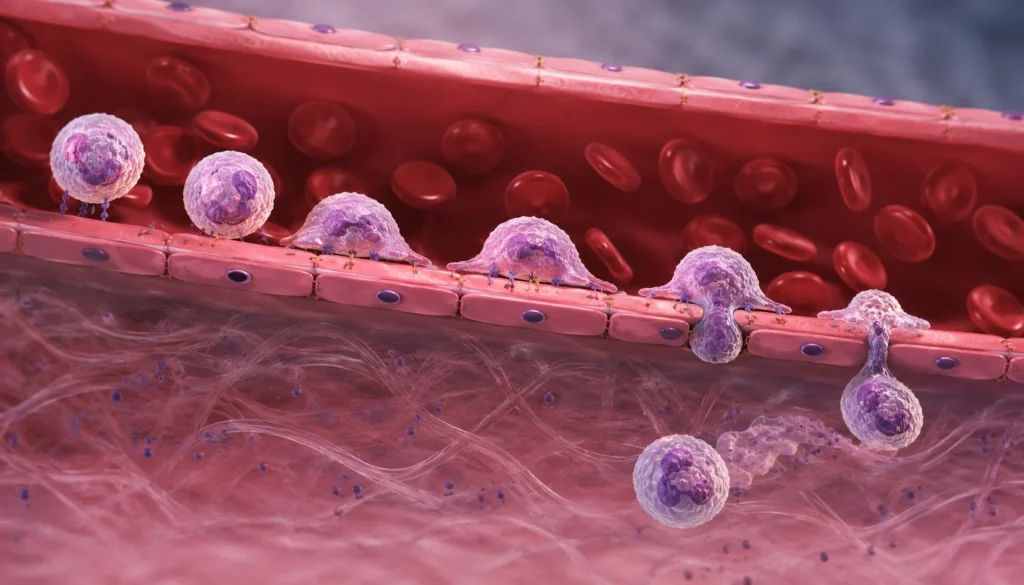
El desplazamiento del neutrófilo desde la circulación sanguínea hacia un tejido lesionado o infectado constituye uno de los procesos más sofisticados de la respuesta inmunitaria innata. Este fenómeno, denominado extravasación leucocitaria o migración transendotelial, permite que los neutrófilos abandonen el compartimento intravascular y alcancen con rapidez el sitio donde se requiere su actividad efectora. Se trata de una secuencia estrictamente ordenada de eventos moleculares y celulares que comprende el anclaje inicial, la rodadura, la adhesión firme, el arrastre o desplazamiento intraluminal y la transmigración a través del endotelio vascular. Cada etapa depende de la activación coordinada de moléculas de adhesión, receptores de quimiocinas, proteínas del citoesqueleto y señales intracelulares que modifican tanto al neutrófilo como a la célula endotelial. La pérdida de cualquiera de estos mecanismos compromete gravemente la capacidad del organismo para combatir infecciones, mientras que su activación excesiva contribuye al desarrollo de inflamación crónica, lesión tisular y enfermedades autoinmunes.
En condiciones fisiológicas, los neutrófilos circulan libremente por el centro del flujo sanguíneo sin establecer contactos estables con el endotelio vascular. Esto ocurre porque el endotelio sano mantiene un estado antiinflamatorio caracterizado por una expresión muy baja de moléculas de adhesión y por la producción continua de óxido nítrico y prostaciclina, sustancias que disminuyen la adhesividad celular y conservan la fluidez del flujo sanguíneo. Además, la elevada velocidad del flujo y las fuerzas de cizallamiento impiden que los neutrófilos permanezcan adheridos a la superficie vascular. Estas características aseguran que los leucocitos únicamente abandonen la circulación cuando exista una señal inflamatoria real.
Cuando ocurre una infección, una lesión traumática o una necrosis tisular, las células residentes del tejido, especialmente macrófagos, mastocitos, células dendríticas y células epiteliales, reconocen patrones moleculares asociados con microorganismos o con daño celular mediante receptores especializados. Como consecuencia liberan grandes cantidades de citocinas proinflamatorias, entre las que destacan el factor de necrosis tumoral α, la interleucina 1β y la interleucina 6, además de quimiocinas como CXCL8 y múltiples mediadores lipídicos derivados del ácido araquidónico. Estas sustancias difunden hacia la microcirculación vecina y activan rápidamente al endotelio vascular, modificando profundamente su fenotipo.
La activación endotelial induce la síntesis y exposición de numerosas moléculas de adhesión que normalmente permanecen ausentes o expresadas en concentraciones muy bajas. Entre las primeras en aparecer se encuentra P-selectina, que es movilizada desde los cuerpos de Weibel-Palade hacia la superficie luminal pocos minutos después del estímulo inflamatorio. Posteriormente aumenta la síntesis de E-selectina, así como de las moléculas de adhesión ICAM-1 y VCAM-1. Paralelamente, las quimiocinas producidas en el tejido son transportadas hasta la superficie luminal del endotelio, donde quedan inmovilizadas mediante su unión a proteoglucanos de heparán sulfato. De esta forma, el endotelio no solamente adquiere capacidad adhesiva, sino que también presenta señales quimiotácticas directamente a los neutrófilos circulantes.
El primer contacto entre el neutrófilo y el endotelio corresponde al anclaje inicial o captura. Este evento consiste en el establecimiento de interacciones extremadamente breves entre las selectinas expresadas por el endotelio y sus ligandos presentes sobre la membrana del neutrófilo. El principal ligando leucocitario es PSGL-1, una glucoproteína intensamente glucosilada que contiene determinantes carbohidratados como sialyl-Lewis X, indispensables para el reconocimiento por P-selectina y E-selectina. Debido a que estas uniones poseen una afinidad relativamente baja y una rápida cinética de asociación y disociación, el neutrófilo no se detiene completamente, sino que reduce parcialmente su velocidad mientras permanece en contacto repetitivo con la superficie endotelial.
La rodadura constituye la consecuencia directa de estas interacciones transitorias. Durante esta fase, el neutrófilo establece y rompe continuamente enlaces con las selectinas endoteliales, lo que genera un movimiento semejante al rodamiento de una esfera sobre una superficie. La velocidad disminuye de cientos de micrómetros por segundo hasta valores considerablemente menores, permitiendo que la célula permanezca suficiente tiempo en contacto con el endotelio para detectar señales quimiotácticas. La rodadura representa un mecanismo altamente eficiente porque posibilita el muestreo continuo del estado inflamatorio del endotelio sin producir obstrucción permanente del flujo sanguíneo.
La importancia funcional de la rodadura radica en que acerca físicamente al neutrófilo a las quimiocinas inmovilizadas sobre la superficie endotelial. Estas moléculas se unen a receptores específicos acoplados a proteínas G, principalmente CXCR1 y CXCR2. La activación de estos receptores desencadena una cascada de señalización intracelular que involucra proteínas G heterotriméricas, fosfolipasa C, fosfatidilinositol 3-cinasa, incremento de calcio citosólico, activación de Rap1, talina y kindlina, culminando en un fenómeno conocido como activación de integrinas mediante señalización desde el interior hacia el exterior. Como resultado, las integrinas presentes en la membrana del neutrófilo experimentan un cambio conformacional que incrementa notablemente su afinidad por los ligandos endoteliales.
La adhesión firme ocurre cuando las integrinas activadas establecen uniones de alta afinidad con las moléculas de adhesión expresadas por el endotelio. Las principales integrinas involucradas son LFA-1 y Mac-1, ambas pertenecientes a la familia β2. LFA-1 reconoce principalmente ICAM-1 e ICAM-2, mientras que Mac-1 interactúa con ICAM-1 y diversos componentes de la matriz extracelular. Estas interacciones poseen una fuerza suficiente para vencer las elevadas fuerzas de cizallamiento generadas por el flujo sanguíneo, produciendo la detención completa del neutrófilo sobre la pared vascular. La adhesión firme constituye el punto de no retorno del proceso migratorio, ya que a partir de este momento el neutrófilo queda comprometido con el abandono de la circulación.
Durante la adhesión firme se producen profundas modificaciones estructurales del neutrófilo. El citoesqueleto de actina experimenta una reorganización dinámica que induce la polarización celular. En el extremo anterior aparece un lamelipodio rico en actina polimerizada, mientras que el extremo posterior desarrolla un uropodio encargado de mantener la tensión celular y facilitar el desplazamiento. Simultáneamente ocurre la redistribución de receptores de membrana, moléculas de adhesión y organelos intracelulares, estableciendo una clara polaridad funcional indispensable para la migración dirigida.
Una vez adherido, el neutrófilo inicia el arrastre o desplazamiento intraluminal sobre la superficie endotelial. En esta fase, la célula no permanece inmóvil, sino que migra lentamente siguiendo gradientes de quimiocinas y explorando el endotelio en busca de sitios anatómicamente favorables para atravesarlo. Este desplazamiento depende principalmente de Mac-1, LFA-1, ICAM-1 y de la reorganización continua del citoesqueleto. El neutrófilo establece sucesivos ciclos de formación y liberación de adhesiones, similares a los observados durante la migración celular en otros tejidos. El objetivo del arrastre consiste en localizar regiones donde las uniones intercelulares presentan mayor susceptibilidad al paso leucocitario, generalmente cerca de las vénulas poscapilares.
La transmigración representa la etapa final del proceso de extravasación. Consiste en el paso físico del neutrófilo desde el compartimento intravascular hacia el espacio intersticial. Existen dos mecanismos principales. En la vía paracelular, la más frecuente, el neutrófilo atraviesa las uniones existentes entre células endoteliales. Para ello, induce una reorganización transitoria de proteínas de unión como VE-cadherina, permitiendo la apertura reversible del espacio intercelular sin comprometer de manera permanente la integridad de la barrera vascular. Este proceso requiere la participación coordinada de PECAM-1, JAM-A, JAM-B, JAM-C, CD99 y ESAM, moléculas expresadas tanto por el endotelio como por el propio neutrófilo.
En un porcentaje menor de casos ocurre la migración transcelular. En este mecanismo, el neutrófilo atraviesa directamente el cuerpo de la célula endotelial mediante la formación de canales transitorios ricos en caveolina, vesículas de transporte y reorganización localizada del citoesqueleto. Aunque inicialmente se consideró un fenómeno excepcional, actualmente se reconoce que puede adquirir mayor relevancia en determinados órganos y durante ciertos procesos inflamatorios.
Después de atravesar el endotelio, el neutrófilo aún debe cruzar la membrana basal vascular. Esta estructura representa una barrera física compuesta principalmente por colágeno tipo IV, laminina, nidógeno y proteoglucanos. El paso ocurre aprovechando regiones fisiológicamente menos densas de la membrana basal y mediante la liberación localizada de proteasas, entre ellas elastasa neutrofílica, metaloproteinasas de matriz y otras enzimas capaces de remodelar transitoriamente la matriz extracelular. Este fenómeno se encuentra estrictamente regulado para minimizar el daño al tejido circundante.
Una vez en el intersticio, el neutrófilo continúa migrando mediante quimiotaxis hacia el foco inflamatorio. El movimiento dirigido depende de gradientes crecientes de quimiocinas, componentes del complemento como C5a, leucotrieno B4, péptidos N-formilados bacterianos y múltiples productos derivados de microorganismos o células lesionadas. El neutrófilo integra simultáneamente estas señales mediante complejas redes de transducción intracelular que regulan la polimerización de actina, la contracción mediada por miosina II y la orientación permanente de la célula hacia el estímulo quimiotáctico dominante.
La secuencia completa de anclaje, rodadura, adhesión, arrastre y transmigración constituye un ejemplo de regulación espacial y temporal extraordinariamente precisa. Cada etapa prepara a la siguiente mediante mecanismos de activación progresiva que incrementan la especificidad del proceso. Las selectinas permiten un reconocimiento inicial reversible; las quimiocinas activan funcionalmente las integrinas; las integrinas estabilizan la adhesión; el arrastre identifica el sitio óptimo de salida, y las moléculas especializadas de las uniones intercelulares facilitan el paso hacia el tejido. Esta organización secuencial evita la extravasación indiscriminada de leucocitos y concentra la respuesta inflamatoria exclusivamente en las zonas donde existe una necesidad biológica real.
La relevancia clínica de este proceso queda demostrada por diversas inmunodeficiencias hereditarias. En la deficiencia de adhesión leucocitaria tipo I, causada por mutaciones del gen ITGB2, las integrinas β2 son incapaces de mediar la adhesión firme, por lo que los neutrófilos permanecen circulando en sangre y no alcanzan los tejidos infectados. En la deficiencia de adhesión leucocitaria tipo II, la alteración de la fucosilación impide la formación de sialyl-Lewis X, comprometiendo la interacción con las selectinas y, por tanto, la rodadura. En la deficiencia de adhesión leucocitaria tipo III, las integrinas están presentes, pero no pueden activarse debido a alteraciones en kindlin-3. En todos estos trastornos predominan infecciones bacterianas recurrentes, retraso en la cicatrización, ausencia de formación adecuada de pus y neutrofilia persistente, demostrando que la extravasación es indispensable para la función protectora de los neutrófilos.
Por el contrario, una activación exagerada de la extravasación neutrofílica contribuye significativamente a la fisiopatología de enfermedades inflamatorias y autoinmunes, incluyendo artritis reumatoide, vasculitis, enfermedad inflamatoria intestinal, síndrome de dificultad respiratoria aguda, isquemia seguida de reperfusión y sepsis. En estas condiciones, la infiltración masiva de neutrófilos favorece la liberación excesiva de especies reactivas de oxígeno, proteasas, citocinas y trampas extracelulares de neutrófilos, produciendo lesión del tejido sano además de la eliminación del agente agresor.


Fuente y lecturas recomendadas:
- Alon, R., & Ley, K. (2008). Cells on the run: Shear-regulated integrin activation in leukocyte rolling and arrest on endothelial cells. Current Opinion in Cell Biology, 20(5), 525-532.
- Carman, C. V., & Springer, T. A. (2004). A transmigratory cup in leukocyte diapedesis both through individual vascular endothelial cells and between them. Journal of Cell Biology, 167(2), 377-388.
- Kolaczkowska, E., & Kubes, P. (2013). Neutrophil recruitment and function in health and inflammation. Nature Reviews Immunology, 13(3), 159-175.
- Ley, K., Laudanna, C., Cybulsky, M. I., & Nourshargh, S. (2007). Getting to the site of inflammation: The leukocyte adhesion cascade updated. Nature Reviews Immunology, 7(9), 678-689.
- Muller, W. A. (2011). Mechanisms of leukocyte transendothelial migration. Annual Review of Pathology: Mechanisms of Disease, 6, 323-344.
- Nourshargh, S., Hordijk, P. L., & Sixt, M. (2010). Breaching multiple barriers: Leukocyte motility through venular walls and the interstitium. Nature Reviews Molecular Cell Biology, 11(5), 366-378.
- Phillipson, M., & Kubes, P. (2011). The neutrophil in vascular inflammation. Nature Medicine, 17(11), 1381-1390.
- Vestweber, D. (2015). How leukocytes cross the vascular endothelium. Nature Reviews Immunology, 15(11), 692-704.
- Weninger, W., Biro, M., & Jain, R. (2014). Leukocyte migration in the interstitial space of non-lymphoid organs. Nature Reviews Immunology, 14(4), 232-246.
Aprende administración paso a paso

ADMINISTRACION DESDE CERO