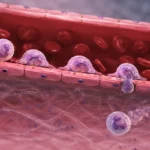
Los neutrófilos constituyen la población más abundante de leucocitos circulantes en la sangre humana y representan la primera línea de defensa del sistema inmunitario innato frente a bacterias, hongos y diversos microorganismos invasores. Su extraordinaria capacidad microbicida depende, en gran medida, de un complejo sistema de organelos citoplasmáticos especializados conocidos como gránulos. Estos compartimentos intracelulares contienen una gran diversidad de enzimas hidrolíticas, proteínas antimicrobianas, proteasas, moléculas reguladoras y componentes necesarios para la generación de especies reactivas del oxígeno. La organización de estos gránulos no es aleatoria, sino que refleja un proceso altamente regulado de diferenciación celular durante la granulopoyesis en la médula ósea, de tal manera que cada tipo de gránulo posee una composición molecular distinta y cumple funciones específicas durante la respuesta inflamatoria.
La formación secuencial de los gránulos ocurre conforme el neutrófilo madura desde el estadio de mieloblasto hasta el neutrófilo segmentado. Los gránulos azurófilos aparecen primero durante la etapa de promielocito; posteriormente se sintetizan los gránulos específicos durante el estadio de mielocito, y finalmente se forman los gránulos de gelatinasa en etapas más avanzadas de maduración. Debido a este desarrollo secuencial, el contenido proteico de cada clase de gránulo refleja el programa de expresión génica predominante en el momento de su formación. Como consecuencia, los tres tipos de gránulos poseen composiciones bioquímicas claramente diferentes y desempeñan funciones complementarias durante la defensa antimicrobiana.
Estos gránulos constituyen reservorios de moléculas proinflamatorias capaces de actuar tanto dentro del fagosoma como en el espacio extracelular. La liberación dirigida de su contenido recibe el nombre de degranulación y constituye uno de los mecanismos efectores más importantes de los neutrófilos. La exocitosis puede producirse de forma parcial, selectiva o masiva, dependiendo de la intensidad del estímulo inflamatorio, del tipo de microorganismo involucrado y del microambiente tisular. Esta regulación permite que los neutrófilos adapten su respuesta según las necesidades del tejido infectado, minimizando, en condiciones fisiológicas, el daño a las células del huésped.
Gránulos azurófilos o primarios
Los gránulos azurófilos representan el tipo de gránulo más antiguo desde el punto de vista evolutivo y también el más rico en enzimas hidrolíticas y proteínas citotóxicas. Se forman durante el estadio de promielocito y presentan características semejantes a los lisosomas clásicos, razón por la cual con frecuencia se consideran lisosomas especializados de los neutrófilos.
Estos gránulos contienen una elevada concentración de enzimas capaces de degradar prácticamente todos los componentes estructurales de bacterias, hongos y células dañadas. Entre sus componentes más importantes se encuentran la mieloperoxidasa, la elastasa de neutrófilos, la catepsina G, la proteinasa 3, la lisozima, la defensina alfa, la azurocidina, diversas catepsinas lisosomales, hidrolasas ácidas, fosfatasas y múltiples proteasas de serina.
La mieloperoxidasa constituye una de las enzimas más características de estos gránulos. Después de la fagocitosis, esta enzima utiliza el peróxido de hidrógeno generado por la nicotinamida adenina dinucleótido fosfato oxidasa para producir ácido hipocloroso mediante la oxidación del ion cloruro. El ácido hipocloroso es uno de los oxidantes biológicos más potentes y destruye proteínas, lípidos, ácidos nucleicos y membranas bacterianas mediante reacciones de halogenación y oxidación, provocando la muerte rápida de numerosos microorganismos.
Las defensinas alfa son pequeños péptidos catiónicos capaces de insertarse en las membranas microbianas debido a su afinidad por los fosfolípidos cargados negativamente. Una vez incorporadas, forman poros que alteran la permeabilidad de la membrana, producen pérdida del gradiente electroquímico y ocasionan la lisis del microorganismo.
La elastasa de neutrófilos, la catepsina G y la proteinasa 3 pertenecen al grupo de las proteasas de serina. Estas enzimas degradan proteínas de la pared bacteriana, factores de virulencia, componentes de la matriz extracelular y proteínas del huésped. Además de ejercer una potente actividad antimicrobiana, participan en la remodelación tisular, la regulación de citocinas y la modulación de la respuesta inflamatoria. Sin embargo, cuando son liberadas de manera excesiva al espacio extracelular pueden producir lesión importante del tejido pulmonar, vascular y conectivo.
La azurocidina posee actividad antimicrobiana directa y también funciona como potente mediador inflamatorio. Incrementa la permeabilidad vascular, favorece el reclutamiento de monocitos y potencia la migración de otros leucocitos hacia el foco infeccioso.
La lisozima hidroliza los enlaces glucosídicos del peptidoglucano presente en la pared bacteriana, especialmente en bacterias Gram positivas, debilitando su estructura y favoreciendo la acción de otras enzimas proteolíticas.
Gránulos específicos o secundarios
Los gránulos específicos aparecen durante la etapa de mielocito y contienen un conjunto diferente de proteínas cuya función principal consiste en facilitar la eliminación microbiana, regular la inflamación y preparar al neutrófilo para la migración hacia los tejidos.
Estos gránulos contienen lactoferrina, lisozima, lipocalina asociada con gelatinasa de neutrófilos, colagenasa tipo IV, proteínas transportadoras, receptores de membrana y componentes del complejo nicotinamida adenina dinucleótido fosfato oxidasa, incluyendo citocromo b558.
La lactoferrina constituye una glicoproteína con elevada afinidad por el hierro férrico. Su principal función consiste en secuestrar hierro libre del medio extracelular. Dado que numerosos microorganismos dependen de este metal para llevar a cabo procesos metabólicos esenciales, la disminución de su disponibilidad limita el crecimiento bacteriano mediante un mecanismo conocido como inmunidad nutricional. Además, la lactoferrina posee actividad bactericida directa, neutraliza lipopolisacáridos bacterianos, inhibe la formación de biopelículas y modula la producción de citocinas inflamatorias.
La lipocalina asociada con gelatinasa de neutrófilos ejerce una función semejante mediante el secuestro de sideróforos bacterianos, moléculas sintetizadas por numerosas bacterias para captar hierro del ambiente. Al impedir que los microorganismos recuperen hierro mediante estos sistemas, limita significativamente su proliferación.
Los gránulos específicos también contienen componentes fundamentales del sistema nicotinamida adenina dinucleótido fosfato oxidasa. Durante la activación del neutrófilo, estas proteínas migran hacia la membrana del fagosoma, donde participan en el ensamblaje del complejo enzimático responsable de producir anión superóxido. A partir de esta molécula se generan múltiples especies reactivas del oxígeno con potente actividad microbicida.
Otra función importante de los gránulos específicos consiste en transportar numerosas proteínas hacia la membrana plasmática del neutrófilo. Entre ellas destacan diversas integrinas, receptores para quimiocinas y moléculas de adhesión que incrementan la capacidad del neutrófilo para adherirse al endotelio vascular, atravesar la pared de los vasos sanguíneos y migrar hacia el tejido inflamado.
Gránulos de gelatinasa o terciarios
Los gránulos terciarios representan el último tipo de gránulo sintetizado durante la maduración del neutrófilo y se especializan principalmente en facilitar la migración celular a través de la matriz extracelular.
Su componente más abundante es la gelatinasa B, también denominada metaloproteinasa de matriz 9. Esta enzima degrada gelatina, colágeno tipo IV, elastina y diversos componentes de la membrana basal. Gracias a esta actividad proteolítica, los neutrófilos pueden atravesar con mayor facilidad la barrera endotelial y desplazarse hacia los tejidos infectados.
Además de la gelatinasa B, estos gránulos contienen otras metaloproteinasas, proteínas reguladoras de la migración celular y receptores que participan en la interacción con la matriz extracelular.
La liberación controlada de gelatinasa contribuye a remodelar el tejido inflamado y facilita el acceso de otras células inmunitarias al sitio de infección. No obstante, cuando esta liberación es excesiva o persistente, puede favorecer la destrucción de cartílago, membrana basal, vasos sanguíneos y otros componentes del tejido conectivo, fenómeno implicado en enfermedades inflamatorias crónicas, artritis reumatoide, enfermedad pulmonar obstructiva crónica, lesión pulmonar aguda y diversas enfermedades cardiovasculares.
Liberación del contenido granular hacia el fagosoma
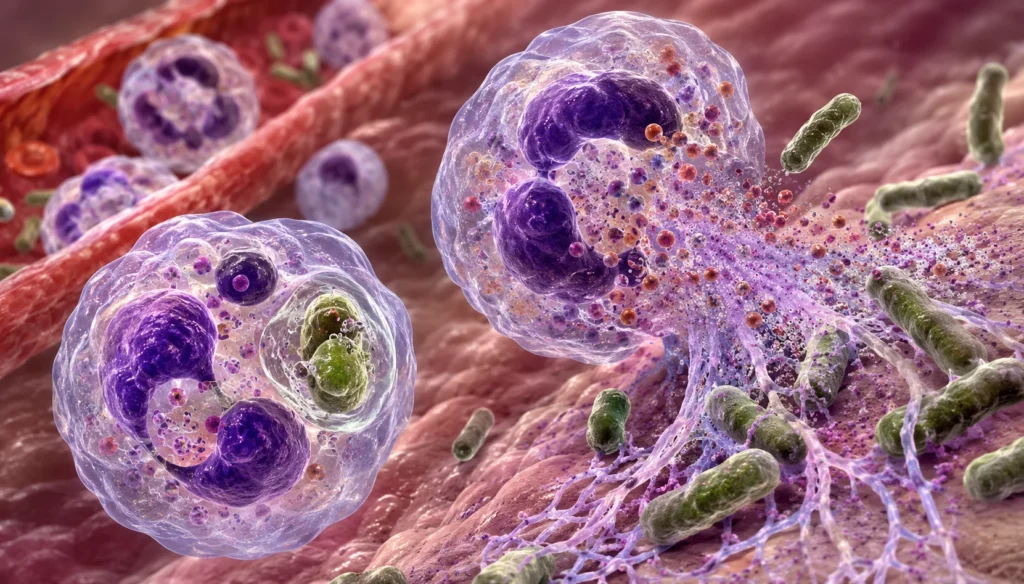
El destino fisiológico más importante del contenido granular es el fagosoma. Cuando un neutrófilo reconoce un microorganismo mediante receptores para inmunoglobulinas, complemento o patrones moleculares asociados con patógenos, la membrana plasmática rodea completamente al agente infeccioso formando una vesícula intracelular denominada fagosoma.
Posteriormente, los diferentes tipos de gránulos se fusionan de manera secuencial con el fagosoma. Inicialmente participan los gránulos específicos y los de gelatinasa, que aportan componentes necesarios para ensamblar el sistema productor de especies reactivas del oxígeno. Después se fusionan los gránulos azurófilos, liberando enzimas lisosomales y proteínas antimicrobianas directamente sobre el microorganismo.
Como consecuencia de esta fusión se forma el fagolisosoma, un compartimento altamente especializado donde coinciden un ambiente ácido, elevadas concentraciones de enzimas proteolíticas, péptidos antimicrobianos y especies reactivas del oxígeno y del cloro. La acción simultánea de estos mecanismos produce una destrucción extremadamente eficiente de bacterias y hongos.
La localización intracelular de estos procesos resulta fundamental para evitar daño innecesario sobre las células vecinas, ya que concentra la actividad citotóxica exclusivamente alrededor del microorganismo fagocitado.
Liberación del contenido granular al medio extracelular
Cuando los microorganismos son demasiado grandes para ser fagocitados, como ocurre con algunos hongos filamentosos, agregados bacterianos o parásitos multicelulares, los neutrófilos pueden liberar directamente el contenido de sus gránulos hacia el espacio extracelular mediante exocitosis.
Durante este proceso, las membranas granulares se fusionan con la membrana plasmática y descargan enzimas proteolíticas, péptidos antimicrobianos y proteínas inflamatorias sobre el tejido circundante.
Esta liberación extracelular permite destruir microorganismos inaccesibles para la fagocitosis, degradar factores de virulencia bacteriana, modificar la matriz extracelular, activar otras células inmunitarias y amplificar la respuesta inflamatoria.
Sin embargo, debido a que muchas de estas enzimas no distinguen entre proteínas microbianas y proteínas del huésped, su liberación excesiva constituye una importante causa de lesión tisular. La elastasa, la catepsina G, la proteinasa 3 y la metaloproteinasa de matriz 9 pueden degradar colágeno, elastina, laminina, fibronectina y otras proteínas estructurales, favoreciendo necrosis tisular, alteraciones vasculares y fibrosis secundaria.
Regulación de la degranulación
La liberación del contenido granular está regulada mediante complejas vías de señalización intracelular que incluyen receptores para inmunoglobulinas, receptores para complemento, receptores tipo Toll, receptores acoplados a proteínas G y diversas cinasas intracelulares.
Existe una jerarquía funcional en la movilización de los gránulos. Los gránulos secretores y los de gelatinasa son los primeros en exocitarse debido a que requieren niveles relativamente bajos de activación celular. Los gránulos específicos necesitan una estimulación intermedia, mientras que los gránulos azurófilos solamente se liberan ante estímulos intensos, debido a que contienen las moléculas potencialmente más citotóxicas.
Este mecanismo escalonado constituye una estrategia de protección del huésped, ya que reduce el riesgo de lesión tisular innecesaria durante respuestas inflamatorias leves y reserva el arsenal proteolítico más agresivo para infecciones graves.
La regulación de este proceso también depende del citoesqueleto, de proteínas Rab, del complejo soluble sensible a N-etilmaleimida y de múltiples proteínas de fusión vesicular que controlan la dirección y el momento exacto de la exocitosis.
Relación con las trampas extracelulares de neutrófilos
Además de la degranulación clásica, numerosos componentes de los gránulos participan en la formación de trampas extracelulares de neutrófilos. Durante este proceso, denominado formación de trampas extracelulares, el neutrófilo libera cromatina descondensada asociada con proteínas granulares, especialmente mieloperoxidasa, elastasa y diversas proteasas.
Estas estructuras forman redes extracelulares que inmovilizan microorganismos y concentran elevadas cantidades de moléculas antimicrobianas alrededor de ellos, incrementando la eficacia de su eliminación.
Aunque este mecanismo resulta beneficioso durante muchas infecciones, una producción excesiva de trampas extracelulares puede favorecer trombosis, daño endotelial, enfermedades autoinmunes y lesión orgánica asociada con inflamación sistémica.
Importancia fisiopatológica
El adecuado funcionamiento de los tres tipos de gránulos resulta indispensable para la inmunidad innata. Alteraciones hereditarias o adquiridas que afectan la formación, el contenido o la liberación de estos gránulos incrementan significativamente la susceptibilidad a infecciones recurrentes.
Por otra parte, la activación excesiva de los neutrófilos y la liberación descontrolada de sus enzimas proteolíticas contribuyen al desarrollo de múltiples enfermedades inflamatorias, entre ellas síndrome de dificultad respiratoria aguda, enfermedad pulmonar obstructiva crónica, fibrosis pulmonar, artritis reumatoide, vasculitis asociadas con anticuerpos anticitoplasma de neutrófilos, aterosclerosis, lesión por isquemia y reperfusión, sepsis y diversas enfermedades autoinmunes.
Los gránulos neutrofílicos representan un sistema efector extraordinariamente eficiente para eliminar microorganismos invasores, pero al mismo tiempo constituyen una importante fuente potencial de daño tisular cuando sus mecanismos reguladores se alteran. El equilibrio entre la actividad antimicrobiana y la limitación del daño colateral determina en gran medida la eficacia y la seguridad de la respuesta inflamatoria aguda.


Fuente y lecturas recomendadas:
- Amulic, B., Cazalet, C., Hayes, G. L., Metzler, K. D., & Zychlinsky, A. (2012). Neutrophil function: From mechanisms to disease. Annual Review of Immunology, 30, 459-489.
- Borregaard, N. (2010). Neutrophils, from marrow to microbes. Immunity, 33(5), 657-670.
- Borregaard, N., & Cowland, J. B. (1997). Granules of the human neutrophilic polymorphonuclear leukocyte. Blood, 89(10), 3503-3521.
- Cowland, J. B., & Borregaard, N. (2016). Granulopoiesis and granules of human neutrophils. Immunological Reviews, 273(1), 11-28.
- Faurschou, M., & Borregaard, N. (2003). Neutrophil granules and secretory vesicles in inflammation. Microbes and Infection, 5(14), 1317-1327.
- Kobayashi, S. D., Malachowa, N., & DeLeo, F. R. (2018). Neutrophils and bacterial immune evasion. Journal of Innate Immunity, 10(5-6), 432-441.
- Nauseef, W. M. (2007). How human neutrophils kill and degrade microbes: An integrated view. Immunological Reviews, 219(1), 88-102.
- Nauseef, W. M., & Borregaard, N. (2014). Neutrophils at work. Nature Immunology, 15(7), 602-611.
- Papayannopoulos, V. (2018). Neutrophil extracellular traps in immunity and disease. Nature Reviews Immunology, 18(2), 134-147.
- Rosales, C. (2020). Neutrophils at the crossroads of innate and adaptive immunity. Journal of Leukocyte Biology, 108(1), 377-396.
Aprende administración paso a paso

ADMINISTRACION DESDE CERO