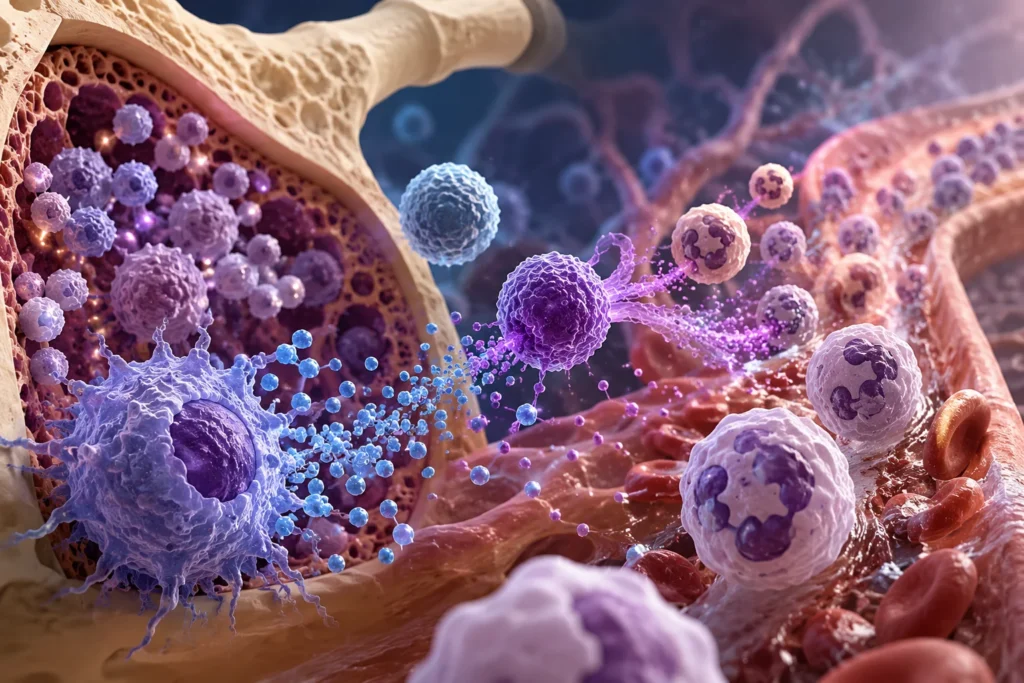
Los neutrófilos constituyen la población más abundante de leucocitos circulantes en el ser humano y representan la primera línea celular de defensa frente a bacterias, hongos y numerosos estímulos inflamatorios. Debido a que poseen una vida media relativamente corta y experimentan un recambio constante, el organismo debe producir cantidades extraordinariamente elevadas de estas células de forma continua. En condiciones fisiológicas, la médula ósea genera aproximadamente entre 10^11 y 10^12 neutrófilos cada día, cifra que puede incrementarse varias veces durante procesos infecciosos o inflamatorios intensos. Esta elevada capacidad de producción permite mantener un número suficiente de neutrófilos maduros en la circulación y, al mismo tiempo, conservar una importante reserva medular capaz de liberarse rápidamente cuando aumenta la demanda. Esta información está sustentada por estudios de hematopoyesis y cinética de neutrófilos que han cuantificado la magnitud del recambio celular diario y la organización de los compartimentos medulares.
La producción continua de neutrófilos recibe el nombre de granulopoyesis. Este proceso ocurre principalmente en la médula ósea y comprende una secuencia ordenada de proliferación, diferenciación y maduración celular que inicia con las células madre hematopoyéticas multipotenciales. Estas células poseen la capacidad de autorrenovarse y de originar todas las líneas celulares sanguíneas. Bajo la influencia de diversos factores de crecimiento y citocinas, una fracción de estas células se diferencia hacia progenitores mieloides comunes y posteriormente hacia progenitores granulocíticos específicos, los cuales originarán la serie neutrofílica. Cada etapa del proceso se encuentra regulada por señales moleculares precisas que determinan si una célula continúa proliferando, inicia su diferenciación o completa su maduración funcional. La evidencia experimental demuestra que esta regulación depende de una compleja interacción entre células hematopoyéticas, células estromales medulares y múltiples citocinas.
Entre todos los factores reguladores de la granulopoyesis, el factor estimulante de las colonias de granulocitos, conocido como G-CSF, constituye el principal regulador fisiológico de la producción de neutrófilos. Se trata de una glucoproteína perteneciente a la familia de los factores estimulantes de colonias cuya función consiste en incrementar la producción, diferenciación, supervivencia y liberación de neutrófilos desde la médula ósea hacia la circulación periférica. Diversos tipos celulares producen G-CSF, incluyendo macrófagos, monocitos, fibroblastos, células endoteliales, células estromales medulares y otros tejidos sometidos a estimulación inflamatoria. Su síntesis aumenta de manera importante durante infecciones bacterianas, lesiones tisulares y procesos inflamatorios sistémicos, permitiendo adaptar rápidamente la producción de neutrófilos a las necesidades del organismo.
El G-CSF ejerce sus efectos mediante la unión a un receptor específico denominado receptor del factor estimulante de las colonias de granulocitos, el cual pertenece a la familia de receptores de citocinas de tipo I. Este receptor se expresa principalmente en progenitores mieloides y en neutrófilos maduros. La unión del G-CSF induce la dimerización del receptor y activa múltiples vías de señalización intracelular, entre ellas las vías Janus kinase/signal transducer and activator of transcription, phosphoinositide 3-kinase/protein kinase B y mitogen-activated protein kinase. La activación coordinada de estas vías modifica la expresión de numerosos genes relacionados con la proliferación celular, la supervivencia, la diferenciación y la función efectora de los neutrófilos.
Uno de los efectos más importantes del G-CSF consiste en estimular la expansión de los progenitores granulocíticos. Durante esta fase aumenta la velocidad de división celular, lo que incrementa considerablemente el número de células precursoras disponibles para completar su diferenciación. Simultáneamente, el G-CSF reduce la apoptosis fisiológica de estas células inmaduras mediante la regulación de proteínas antiapoptóticas, prolongando así su supervivencia y favoreciendo una mayor producción final de neutrófilos maduros. La evidencia obtenida en modelos animales y estudios clínicos demuestra que la ausencia de señalización por G-CSF ocasiona neutropenia grave, mientras que su administración exógena incrementa rápidamente el número absoluto de neutrófilos circulantes.
Además de estimular la proliferación, el G-CSF acelera la maduración de la serie granulocítica. Durante este proceso, los mieloblastos evolucionan sucesivamente hacia promielocitos, mielocitos, metamielocitos, neutrófilos en banda y finalmente neutrófilos segmentados completamente funcionales. En cada una de estas etapas ocurre una reorganización profunda del programa de expresión génica, la síntesis de gránulos citoplasmáticos especializados, la segmentación progresiva del núcleo y el desarrollo de la maquinaria necesaria para la quimiotaxis, la fagocitosis, la producción de especies reactivas del oxígeno y la formación de trampas extracelulares de neutrófilos. El G-CSF coordina estos procesos regulando factores de transcripción esenciales para la diferenciación mieloide, entre ellos CCAAT/enhancer-binding protein alpha y CCAAT/enhancer-binding protein epsilon.
El G-CSF también facilita la movilización de neutrófilos desde la médula ósea hacia la sangre periférica. En condiciones normales, la médula ósea mantiene un gran reservorio de neutrófilos maduros que supera ampliamente el número presente en la circulación. Cuando aumenta la concentración de G-CSF, disminuye la expresión de moléculas responsables de la retención medular, particularmente el eje formado por C-X-C motif chemokine ligand 12 y su receptor C-X-C chemokine receptor type 4. Como consecuencia, los neutrófilos abandonan la médula ósea con mayor rapidez y pasan al torrente sanguíneo para dirigirse hacia los tejidos inflamados. Este mecanismo explica el aumento rápido del número de neutrófilos observado pocas horas después de la administración terapéutica de G-CSF o durante infecciones bacterianas agudas.
La regulación fisiológica del G-CSF depende de una red compleja de citocinas proinflamatorias. Entre ellas, la interleucina-17 ocupa una posición central como estimulador de la granulopoyesis. La IL-17 corresponde a una familia de citocinas, siendo IL-17A la principal molécula implicada en este proceso. Esta citocina es producida predominantemente por los linfocitos T helper 17, aunque también puede sintetizarse por otras poblaciones celulares, incluyendo linfocitos T gamma delta, células linfoides innatas del grupo 3, células natural killer y algunos neutrófilos activados.
La diferenciación de los linfocitos T hacia el fenotipo T helper 17 requiere un microambiente inflamatorio caracterizado por la presencia de diversas citocinas, entre ellas transforming growth factor beta, IL-6, IL-1 beta e IL-23. Una vez diferenciados, estos linfocitos producen grandes cantidades de IL-17, la cual actúa sobre fibroblastos, células endoteliales, células epiteliales y células estromales de la médula ósea. Estas células responden aumentando la síntesis de G-CSF y de diversas quimiocinas capaces de atraer neutrófilos hacia los focos inflamatorios.
La IL-17 no estimula directamente la proliferación de los progenitores granulocíticos. Su principal función consiste en inducir la producción secundaria de G-CSF por múltiples tejidos. Este mecanismo amplifica considerablemente la granulopoyesis, ya que una pequeña cantidad inicial de IL-17 puede desencadenar una producción elevada de G-CSF y, en consecuencia, una expansión masiva de la línea neutrofílica. La importancia fisiológica de esta vía ha sido demostrada mediante modelos experimentales en los cuales la neutralización de IL-17 disminuye significativamente la producción de G-CSF y reduce la neutrofilia inducida por infecciones bacterianas.
Además de estimular la producción de G-CSF, la IL-17 incrementa la síntesis de diversas quimiocinas, como C-X-C motif chemokine ligand 1, C-X-C motif chemokine ligand 2, C-X-C motif chemokine ligand 5 y C-X-C motif chemokine ligand 8, que favorecen el reclutamiento de neutrófilos hacia los tejidos lesionados. De esta manera, la IL-17 coordina simultáneamente el incremento de la producción medular y la migración de neutrófilos hacia los sitios donde son necesarios, integrando la respuesta hematopoyética con la respuesta inflamatoria periférica.
La producción de IL-17 depende, a su vez, de la acción de la IL-23. Esta citocina pertenece a la familia de la IL-12 y está formada por dos subunidades denominadas p19 y p40. Los principales productores de IL-23 son los macrófagos activados, las células dendríticas y otras células presentadoras de antígenos que reconocen microorganismos mediante receptores de reconocimiento de patrones, incluyendo los receptores tipo Toll.
Cuando un macrófago detecta componentes bacterianos, fúngicos o productos derivados del daño tisular, activa vías intracelulares que culminan con la síntesis de IL-23. Esta citocina actúa sobre los linfocitos T helper 17 ya diferenciados, promoviendo su expansión clonal, aumentando su supervivencia y potenciando la producción sostenida de IL-17. En consecuencia, la IL-23 constituye un regulador indispensable para mantener respuestas prolongadas mediadas por IL-17 durante infecciones persistentes o inflamaciones crónicas.
La relación funcional entre macrófagos, IL-23, linfocitos T helper 17, IL-17 y G-CSF conforma un eje inmunológico conocido como eje IL-23/IL-17/G-CSF. Este circuito representa uno de los mecanismos más importantes mediante los cuales el sistema inmunitario adapta la producción de neutrófilos a la intensidad de la agresión infecciosa. Cuando los macrófagos identifican microorganismos, producen IL-23. La IL-23 estimula los linfocitos T helper 17 para incrementar la síntesis de IL-17. Posteriormente, la IL-17 induce la producción de G-CSF en diversos tejidos, y el G-CSF estimula la granulopoyesis y la liberación de neutrófilos desde la médula ósea. Este sistema permite transformar una señal inmunitaria localizada en una respuesta hematopoyética sistémica altamente eficiente.
Durante infecciones bacterianas graves, este eje puede incrementar varias veces la velocidad normal de producción de neutrófilos. La médula ósea responde expandiendo rápidamente el compartimento proliferativo de progenitores granulocíticos y acelerando la maduración celular. Si la demanda supera la capacidad habitual de producción, incluso pueden liberarse formas menos maduras, como los neutrófilos en banda, fenómeno conocido como desviación hacia la izquierda, el cual constituye un marcador clásico de respuesta inflamatoria intensa.
La regulación de este sistema también depende de mecanismos de retroalimentación negativa que evitan una producción excesiva de neutrófilos. Conforme disminuye la carga microbiana y se resuelve la inflamación, descienden las concentraciones de IL-23, IL-17 y G-CSF. Paralelamente, los neutrófilos envejecidos que regresan a la médula ósea o son fagocitados por macrófagos contribuyen indirectamente a reducir la producción de IL-23 y G-CSF, restaurando gradualmente la hematopoyesis basal. Este mecanismo homeostático impide el desarrollo de neutrofilia persistente y limita el daño tisular asociado con una inflamación excesiva.
La relevancia clínica de esta vía reguladora queda demostrada por diversas enfermedades humanas. Las mutaciones que afectan el receptor del G-CSF pueden producir neutropenia congénita grave debido a una alteración de la granulopoyesis. Asimismo, los tratamientos con G-CSF recombinante constituyen una estrategia terapéutica ampliamente utilizada para prevenir o corregir la neutropenia inducida por quimioterapia, trasplante de médula ósea y otras enfermedades hematológicas. Por otra parte, la activación persistente del eje IL-23/IL-17 contribuye a la fisiopatología de diversas enfermedades inflamatorias y autoinmunitarias, incluyendo psoriasis, espondiloartritis y enfermedad inflamatoria intestinal, en las cuales el exceso de neutrófilos participa en el daño tisular.
La producción continua de neutrófilos representa un proceso dinámico y estrictamente regulado que integra mecanismos hematopoyéticos e inmunológicos. El G-CSF constituye el principal factor efector que controla la proliferación, diferenciación, supervivencia y movilización de los neutrófilos. La IL-17 actúa como un regulador superior que induce la síntesis de G-CSF en respuesta a estímulos inflamatorios, mientras que la IL-23 producida por macrófagos mantiene la actividad de los linfocitos productores de IL-17. La coordinación de estas tres citocinas permite que la médula ósea adapte continuamente la producción de neutrófilos a las necesidades del organismo, garantizando una respuesta rápida y eficaz frente a infecciones y preservando, al mismo tiempo, la homeostasis del sistema inmunitario.


Fuente y lecturas recomendadas:
- Basu, S., Hodgson, G., Katz, M., & Dunn, A. R. (2002). Evaluation of role of G-CSF in the production, survival, and release of neutrophils from bone marrow into circulation. Blood, 100(3), 854–861. https://doi.org/10.1182/blood.V100.3.854
- Borregaard, N. (2010). Neutrophils, from marrow to microbes. Immunity, 33(5), 657–670. https://doi.org/10.1016/j.immuni.2010.11.011
- Craig, R. W., & Foote, J. (2009). Biology of granulocyte colony-stimulating factor. Current Hematology Reports, 8, 183–189.
- Kolls, J. K., & Lindén, A. (2004). Interleukin-17 family members and inflammation. Immunity, 21(4), 467–476. https://doi.org/10.1016/j.immuni.2004.08.018
- Manz, M. G., & Boettcher, S. (2014). Emergency granulopoiesis. Nature Reviews Immunology, 14(5), 302–314. https://doi.org/10.1038/nri3660
- Miossec, P., Korn, T., & Kuchroo, V. K. (2009). Interleukin-17 and type 17 helper T cells. The New England Journal of Medicine, 361(9), 888–898. https://doi.org/10.1056/NEJMra0707449
- Panopoulos, A. D., & Watowich, S. S. (2008). Granulocyte colony-stimulating factor: Molecular mechanisms of action during steady state and emergency hematopoiesis. Cytokine, 42(3), 277–288. https://doi.org/10.1016/j.cyto.2008.03.002
- Stark, M. A., Huo, Y., Burcin, T. L., Morris, M. A., Olson, T. S., & Ley, K. (2005). Phagocytosis of apoptotic neutrophils regulates granulopoiesis via IL-23 and IL-17. Immunity, 22(3), 285–294. https://doi.org/10.1016/j.immuni.2005.01.011
- Takizawa, H., Boettcher, S., & Manz, M. G. (2012). Demand-adapted regulation of early hematopoiesis in infection and inflammation. Blood, 119(13), 2991–3002. https://doi.org/10.1182/blood-2011-12-380113
- Zhu, J., Yamane, H., & Paul, W. E. (2010). Differentiation of effector CD4 T cell populations. Annual Review of Immunology, 28, 445–489. https://doi.org/10.1146/annurev-immunol-030409-101212
Aprende administración paso a paso

ADMINISTRACION DESDE CERO