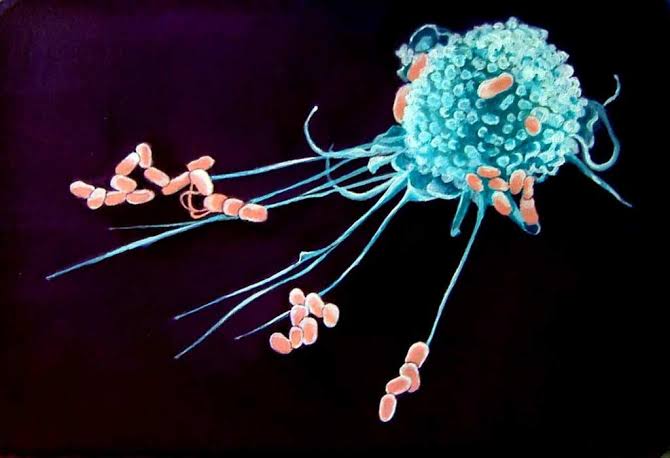
Los macrófagos constituyen una población celular fundamental del sistema inmunitario innato y derivan de la línea hematopoyética mononuclear. Su origen se sitúa en la médula ósea, donde las células madre hematopoyéticas generan monocitos circulantes que posteriormente migran hacia los tejidos periféricos, proceso descrito como parte del sistema fagocítico mononuclear en estudios clásicos de hematología e inmunología celular. La diferenciación de monocitos a macrófagos tisulares ocurre bajo la influencia del microambiente local, que incluye citocinas como el factor estimulante de colonias de macrófagos y señales derivadas del daño tisular o de patógenos. Este proceso ha sido ampliamente caracterizado en la literatura de inmunobiología contemporánea, donde se establece que la plasticidad del linaje mononuclear permite la adaptación funcional del macrófago al tejido residente y al contexto inflamatorio.
Los macrófagos del tejido conjuntivo, históricamente denominados histiocitos, representan la forma residente de estas células en el estroma conectivo. La transición de monocito a macrófago implica cambios morfológicos y funcionales profundos, incluyendo el aumento del volumen citoplasmático, la expansión del sistema lisosomal y la adquisición de capacidades fagocíticas avanzadas. Esta diferenciación ha sido documentada mediante estudios histológicos y de microscopía electrónica que demuestran la reorganización del citoesqueleto y el incremento de organelos implicados en degradación intracelular.
Desde el punto de vista morfológico, los macrófagos tisulares presentan un núcleo excéntrico con forma arriñonada o hendido, rasgo característico asociado a su estado de activación y diferenciación. El citoplasma es abundante y contiene numerosas vesículas lisosomales, esenciales para la degradación de material fagocitado. La membrana plasmática exhibe múltiples prolongaciones digitiformes y pliegues irregulares que incrementan la superficie de contacto con el entorno extracelular, facilitando la captación de partículas mediante mecanismos de endocitosis y fagocitosis. Estas características ultrastructurales han sido descritas con precisión mediante microscopía electrónica de transmisión, la cual ha permitido correlacionar la arquitectura celular con su función inmunitaria.
Los lisosomas constituyen uno de los componentes más abundantes del macrófago y desempeñan un papel central en la digestión intracelular. Estas organelas contienen hidrolasas ácidas capaces de degradar proteínas, lípidos, polisacáridos y ácidos nucleicos, lo cual permite la destrucción eficiente de microorganismos internalizados y detritos celulares. El proceso de fagocitosis culmina en la formación de fagolisosomas, resultado de la fusión entre fagosomas y lisosomas, donde ocurre la degradación enzimática del material ingerido. Este mecanismo ha sido ampliamente documentado en estudios bioquímicos y celulares que demuestran la importancia del ambiente ácido intralisosomal para la activación enzimática.
Además de la digestión intracelular, los macrófagos presentan una notable capacidad secretora. El retículo endoplásmico rugoso y el aparato de Golgi se encuentran altamente desarrollados, lo que refleja una intensa síntesis de proteínas implicadas en la respuesta inmunitaria. Entre estas proteínas se incluyen citocinas proinflamatorias, quimiocinas, factores de crecimiento y enzimas proteolíticas. La secreción ocurre mediante mecanismos de exocitosis constitutiva y regulada, dependiendo del estímulo microambiental. Estudios inmunológicos han demostrado que la activación de receptores de reconocimiento de patrones desencadena la liberación de mediadores inflamatorios esenciales para la coordinación de la respuesta inmunitaria.
Los macrófagos también desempeñan una función esencial como células presentadoras de antígeno. Expresan moléculas del complejo mayor de histocompatibilidad de clase II, las cuales permiten la presentación de péptidos derivados de patógenos a linfocitos T cooperadores CD4 positivo. Este proceso implica la internalización del antígeno, su procesamiento proteolítico en compartimentos endosómicos y su posterior carga en moléculas del complejo mayor de histocompatibilidad II para su exposición en la superficie celular. La interacción entre el macrófago y el linfocito T desencadena la activación de la respuesta inmunitaria adaptativa, un fenómeno ampliamente descrito en la inmunología moderna como un punto de convergencia entre la inmunidad innata y adaptativa.
En el contexto de la inflamación aguda, los neutrófilos son las primeras células en migrar hacia el sitio de lesión tisular, donde realizan fagocitosis y liberación de especies reactivas de oxígeno y enzimas proteolíticas. Este proceso genera un microambiente rico en detritos celulares y mediadores inflamatorios. Posteriormente, los monocitos extravasan desde el torrente sanguíneo hacia el tejido lesionado, donde se diferencian en macrófagos. Este reclutamiento tardío ha sido descrito como una fase de resolución y amplificación de la respuesta inflamatoria, en la cual los macrófagos eliminan restos celulares y microorganismos resistentes a la acción neutrofílica.

Durante la activación macrófaga, se reconocen diferentes estados funcionales, entre los cuales destacan los macrófagos de activación clásica y los de activación alternativa. Los macrófagos de activación clásica, también denominados M1, se inducen principalmente por interferón gamma, factor de necrosis tumoral alfa y lipopolisacárido bacteriano. Este fenotipo se caracteriza por la producción de óxido nítrico mediante la enzima óxido nítrico sintasa inducible, así como por la liberación de citocinas proinflamatorias como interleucina doce y factor de necrosis tumoral alfa. Estas moléculas promueven la destrucción microbiana y amplifican la respuesta inflamatoria local. Estudios experimentales han demostrado que este perfil funcional es crucial para la eliminación de patógenos intracelulares, aunque su activación prolongada puede contribuir al daño tisular crónico.
En contraste, los macrófagos de activación alternativa, denominados M2, se inducen por citocinas como interleucina cuatro e interleucina trece. Este fenotipo está asociado con la resolución de la inflamación, la reparación tisular y la remodelación de la matriz extracelular. Los macrófagos M2 secretan factores como factor de crecimiento endotelial vascular, que promueve la angiogénesis, así como componentes de matriz extracelular que contribuyen a la restauración estructural del tejido. Investigaciones en biología de la reparación tisular han demostrado que estos macrófagos desempeñan un papel central en la transición desde la inflamación hacia la regeneración.
La plasticidad funcional de los macrófagos incluye también la capacidad de fusionarse en condiciones específicas para formar células gigantes multinucleadas. Este fenómeno ocurre especialmente en presencia de cuerpos extraños persistentes y se observa en granulomas. Es importante distinguir entre células gigantes de cuerpo extraño y células gigantes tipo Langhans, estas últimas asociadas principalmente a procesos granulomatosos inmunitarios como la tuberculosis. Ambas formas representan estrategias adaptativas para contener material no degradable.

Fuente y lecturas recomendadas:
- Abbas, A. K., Lichtman, A. H., & Pillai, S. (2021). Cellular and Molecular Immunology (10th ed.). Elsevier.
- Murphy, K., & Weaver, C. (2016). Janeway’s Immunobiology (9th ed.). Garland Science.
- Ross, M. H., & Pawlina, W. (2020). Histology: A Text and Atlas (8th ed.). Wolters Kluwer.
- Alberts, B. et al. (2015). Molecular Biology of the Cell (6th ed.). Garland Science.
- Nathan, C. (2008). Macrophages: multifunctional effector cells. Nature Reviews Immunology, 8, 11–20.
- Unanue, E. R. (2007). Antigen presentation in the immune response. Annual Review of Immunology, 25, 1–34.
- Soehnlein, O., & Lindbom, L. (2010). Phagocyte partnership during the onset and resolution of inflammation. Nature Reviews Immunology, 10, 427–439.
- Murray, P. J., & Wynn, T. A. (2011). Protective and pathogenic functions of macrophage subsets. Nature Reviews Immunology, 11, 723–737.
- Gordon, S., & Martinez, F. O. (2010). Alternative activation of macrophages: mechanism and functions. Immunity, 32, 593–604.
- Wynn, T. A., Chawla, A., & Pollard, J. W. (2013). Macrophage biology in development, homeostasis and disease. Nature, 496, 445–455.
- Ehlers, S., & Schaible, U. E. (2013). The granuloma in tuberculosis: dynamics and function. Current Opinion in Microbiology, 16, 38–44.
Aprende administración paso a paso

ADMINISTRACION DESDE CERO