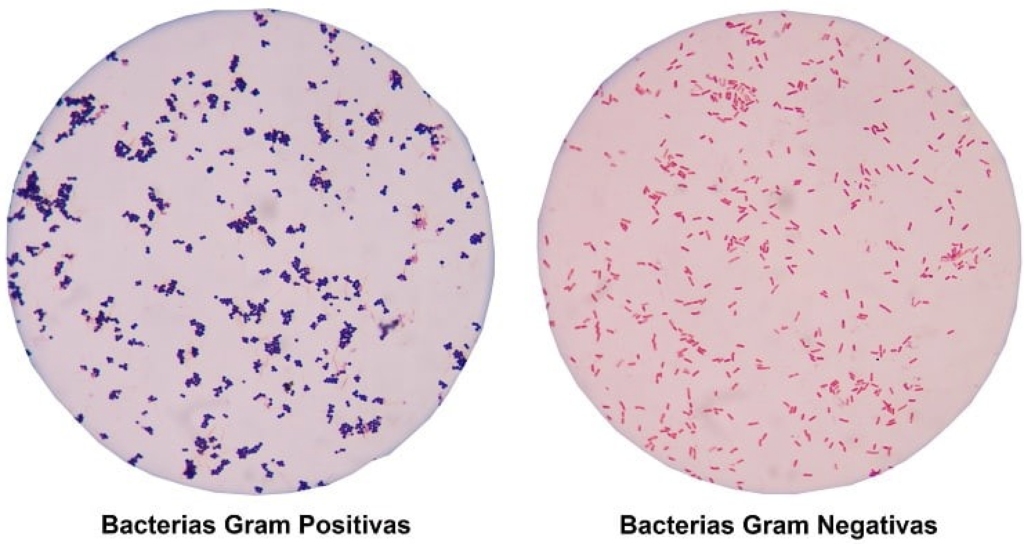
Las bacterias Gram positivas y Gram negativas representan dos grandes grupos de microorganismos procariotas cuya diferenciación se fundamenta principalmente en la arquitectura y composición de su pared celular. Esta estructura no solo determina el resultado de la tinción de Gram, sino que condiciona aspectos esenciales de la biología bacteriana, como la resistencia mecánica, la supervivencia frente a cambios osmóticos, la interacción con el hospedador, la virulencia y la sensibilidad a enzimas y antibióticos. La pared celular, por tanto, debe entenderse como un sistema dinámico y multifuncional, y no únicamente como una envoltura pasiva.
Bacterias Gram positivas
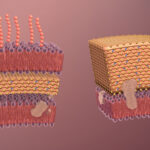
Las bacterias Gram positivas se caracterizan por poseer una pared celular gruesa y multilaminar, compuesta fundamentalmente por peptidoglucano, con un espesor aproximado de entre ciento cincuenta y quinientos angstroms. Esta estructura rodea directamente a la membrana citoplasmática y constituye el componente predominante de la envoltura celular. El peptidoglucano forma una red tridimensional altamente organizada que actúa como un exoesqueleto rígido, comparable funcionalmente al exoesqueleto de los insectos, en el sentido de que proporciona soporte mecánico, define la forma celular y protege frente a fuerzas externas. Sin embargo, a diferencia de los exoesqueletos macroscópicos, esta malla molecular es suficientemente porosa como para permitir la difusión de metabolitos, iones y pequeñas moléculas hasta la membrana plasmática, donde se localizan los sistemas de transporte y de generación de energía.
A nivel molecular, el peptidoglucano de las bacterias Gram positivas está constituido por largas cadenas de glicanos formadas por la alternancia repetitiva de N-acetilglucosamina y ácido N-acetilmurámico. Estas cadenas se proyectan hacia el exterior desde la membrana citoplasmática como si fueran cerdas rígidas y se encuentran entrecruzadas por cortas cadenas peptídicas, lo que confiere a la pared celular una gran resistencia tensional. Esta estructura es esencial no solo para mantener la integridad física de la célula, sino también para procesos fundamentales como el crecimiento y la división bacteriana. Durante la replicación, el peptidoglucano debe ser parcialmente degradado y vuelto a sintetizar de manera controlada, lo que pone de manifiesto su carácter dinámico.
La importancia vital del peptidoglucano se evidencia en su vulnerabilidad frente a la acción de la lisozima. Esta enzima, presente de forma natural en secreciones humanas como las lágrimas y el moco, así como producida por diversos organismos, cataliza la ruptura de los enlaces glucosídicos de la cadena de glicanos. Cuando el peptidoglucano es degradado, la bacteria pierde su principal elemento de contención frente a la elevada presión osmótica interna. Como consecuencia, la membrana citoplasmática no puede soportar el gradiente osmótico y la célula sufre lisis. La eliminación completa de la pared celular da lugar a un protoplasto, una forma extremadamente frágil que solo puede sobrevivir en condiciones artificiales de estabilidad osmótica.
Además del peptidoglucano, la pared celular de las bacterias Gram positivas suele contener una serie de componentes accesorios que cumplen funciones estructurales, fisiológicas e inmunológicas. Entre ellos destacan las proteínas de superficie, los ácidos teicoicos, los ácidos lipoteicoicos y polisacáridos complejos conocidos como polisacáridos C. Muchas proteínas con relevancia patogénica se encuentran unidas de manera covalente al peptidoglucano. Ejemplos clásicos incluyen la proteína M de los estreptococos y la proteína A de Staphylococcus aureus, las cuales contribuyen de manera decisiva a la evasión del sistema inmunitario y a la adhesión a células humanas. Otras proteínas asociadas a la pared celular facilitan la colonización de tejidos y el establecimiento de infecciones.
Los ácidos teicoicos son polímeros aniónicos hidrosolubles formados por unidades repetitivas de fosfatos de polioles, como el glicerol o el ribitol, y se encuentran covalentemente enlazados al peptidoglucano. Estos compuestos son esenciales para la viabilidad celular, ya que contribuyen a la estabilidad estructural de la pared, regulan la carga superficial de la bacteria y participan en la homeostasis iónica. Los ácidos lipoteicoicos, estrechamente relacionados, poseen además un componente lipídico que los ancla a la membrana citoplasmática. Ambos tipos de ácidos actúan como importantes antígenos de superficie, permitiendo la diferenciación de serotipos bacterianos y facilitando la adhesión tanto a otras bacterias como a receptores específicos en las células del hospedador.
Desde el punto de vista de la patogenicidad, los ácidos teicoicos y lipoteicoicos desempeñan un papel relevante como factores de virulencia. Los ácidos lipoteicoicos pueden liberarse al medio o al interior del hospedador, donde interactúan con receptores de reconocimiento de patrones del sistema inmunitario innato. Aunque su potencia inflamatoria es menor que la del lipopolisacárido de las bacterias Gram negativas, estos compuestos son capaces de activar respuestas inmunitarias protectoras, contribuyendo a la inflamación local y sistémica.
Bacterias Gram negativas
Las bacterias Gram negativas se distinguen de las Gram positivas por poseer una envoltura celular notablemente más compleja, tanto desde el punto de vista estructural como químico. Esta complejidad no es un detalle accesorio, sino el resultado de una organización altamente especializada que condiciona su fisiología, su interacción con el entorno, su capacidad patogénica y su comportamiento frente al sistema inmunitario del hospedador y frente a los antimicrobianos. En las bacterias Gram negativas, la pared celular no puede entenderse como una única capa rígida, sino como un sistema multicapa integrado por la membrana citoplasmática, una capa delgada de peptidoglucano, el espacio periplásmico y una membrana externa exclusiva de este grupo bacteriano.
Inmediatamente por fuera de la membrana citoplasmática se localiza una capa fina de peptidoglucano, que representa únicamente entre un cinco y un diez por ciento del peso total de la pared celular Gram negativa. A diferencia de lo observado en las bacterias Gram positivas, este peptidoglucano no forma una estructura gruesa ni multilaminar, sino una malla relativamente delgada cuya función principal es mantener la forma celular y aportar un nivel básico de rigidez mecánica. En este grupo bacteriano, el peptidoglucano no es el componente dominante de la protección celular, y su papel estructural se ve complementado y reforzado por la presencia de la membrana externa. Es importante destacar que en las bacterias Gram negativas no existen ácidos teicoicos ni lipoteicoicos, lo que marca una diferencia química fundamental con las Gram positivas.
Entre la membrana citoplasmática interna y la membrana externa se encuentra el espacio periplásmico, un compartimento funcional claramente definido que no debe considerarse un simple espacio vacío. El periplasma contiene una gran variedad de proteínas y enzimas esenciales para la supervivencia bacteriana. En este compartimento se localizan componentes de sistemas de transporte implicados en la captación de hierro, azúcares, aminoácidos y otros metabolitos, así como proteínas que participan en la exportación de sustancias hacia el exterior. Además, el periplasma alberga numerosas enzimas hidrolíticas, como proteasas, fosfatasas, lipasas, nucleasas y enzimas degradadoras de carbohidratos, cuya función es fragmentar macromoléculas complejas en unidades más pequeñas que puedan ser transportadas al citoplasma para su metabolismo.
En las especies Gram negativas patógenas, el espacio periplásmico adquiere una importancia adicional, ya que en él se concentran muchos factores de virulencia. Enzimas como colagenasas, hialuronidasas y proteasas contribuyen a la degradación de tejidos del hospedador, facilitando la invasión y la diseminación bacteriana. Asimismo, la presencia de β-lactamasas en el periplasma explica uno de los mecanismos más relevantes de resistencia a antibióticos, ya que estas enzimas pueden inactivar antibióticos β-lactámicos antes de que alcancen sus dianas en la síntesis del peptidoglucano.
La envoltura celular Gram negativa está atravesada por múltiples sistemas de transporte que regulan de manera estricta la entrada y salida de moléculas. Entre estos sistemas destacan los dispositivos de secreción de tipo uno a tipo cinco, complejos multiproteicos que permiten el paso dirigido de proteínas a través de una o ambas membranas. La producción de estos sistemas suele inducirse durante la infección y está estrechamente asociada con la virulencia. En particular, el sistema de secreción de tipo tres constituye un factor de virulencia mayor en diversas bacterias patógenas. Este sistema posee una estructura compleja que atraviesa tanto la membrana interna como la externa y funciona de manera análoga a una jeringa molecular, inyectando proteínas bacterianas directamente en células eucariotas o incluso en otras bacterias. Las proteínas transferidas mediante este mecanismo alteran funciones celulares del hospedador, favorecen la adhesión, bloquean respuestas inmunitarias o promueven la supervivencia intracelular del microorganismo.
Una característica definitoria y exclusiva de las bacterias Gram negativas es la presencia de la membrana externa, una estructura que rodea a la célula como una envoltura rígida comparable a un saco resistente. Esta membrana cumple un papel crucial en el mantenimiento de la forma bacteriana y actúa como una potente barrera de permeabilidad. Gracias a ella, la bacteria queda protegida frente a moléculas grandes, como proteínas antimicrobianas y enzimas como la lisozima, así como frente a moléculas hidrofóbicas, entre ellas diversos agentes antimicrobianos. Esta función de barrera resulta especialmente importante para bacterias que habitan entornos hostiles, como el tracto digestivo, donde deben resistir la acción de sales biliares, enzimas digestivas y cambios de pH, como ocurre en muchas enterobacterias.
Desde el punto de vista químico, la membrana externa presenta una estructura de bicapa asimétrica única entre las membranas biológicas. La cara interna está compuesta por fosfolípidos similares a los presentes en otras membranas bacterianas, mientras que la cara externa está formada casi exclusivamente por lipopolisacárido. Salvo durante su síntesis y transporte, el lipopolisacárido se localiza únicamente en esta región de la célula, lo que subraya su especificidad estructural.
El lipopolisacárido, también conocido como endotoxina, es una de las moléculas biológicamente más activas producidas por bacterias. Actúa como un patrón molecular asociado a patógenos y es reconocido por receptores del sistema inmunitario innato del hospedador. El lipopolisacárido puede desprenderse de la superficie bacteriana y difundirse en el organismo del hospedador, donde se une a receptores presentes en células como macrófagos, células dendríticas y linfocitos B. Esta interacción desencadena cascadas de señalización que conducen a la producción de citocinas proinflamatorias, la activación de la respuesta inmune y la inducción de fiebre.
Cuando grandes cantidades de endotoxina alcanzan el torrente sanguíneo, pueden producirse efectos sistémicos graves, como inflamación descontrolada, shock séptico y la reacción de Shwartzman, caracterizada por coagulación intravascular diseminada. Algunas bacterias, como las del género Neisseria, liberan grandes cantidades de una forma relacionada y truncada del lipopolisacárido denominada lipooligosacárido, cuya liberación masiva se asocia con fiebre elevada y síntomas clínicos severos.
Aunque el repertorio de proteínas presentes en la membrana es relativamente limitado en comparación con otros compartimentos celulares, varias de estas proteínas se encuentran en concentraciones muy elevadas. Como resultado, el contenido proteico total de la membrana externa supera al de la membrana citoplasmática, lo que subraya su papel central en la fisiología bacteriana, en la interacción con el entorno y en la resistencia frente a agentes antimicrobianos.
Una característica clave de muchas proteínas de la membrana externa es su naturaleza transmembrana. Estas proteínas atraviesan completamente la bicapa lipídica y adoptan conformaciones estructurales altamente conservadas, adaptadas a la arquitectura particular de esta membrana. Entre ellas destaca un grupo funcionalmente esencial conocido como porinas. Las porinas forman canales acuosos que permiten la difusión pasiva de moléculas hidrofílicas de bajo peso molecular, generalmente inferiores a setecientos daltones. Gracias a estas estructuras, la bacteria puede captar nutrientes esenciales, como azúcares simples, aminoácidos y ciertos iones, sin comprometer la integridad de la barrera protectora que representa la membrana externa.
La selectividad de las porinas es un elemento crítico para la supervivencia bacteriana. Aunque permiten el paso de pequeñas moléculas hidrofílicas, restringen de manera eficaz la entrada de moléculas grandes y de compuestos hidrofóbicos, entre ellos muchos antibióticos. Esta función de filtrado contribuye de forma significativa a la resistencia intrínseca de las bacterias Gram negativas frente a múltiples agentes antimicrobianos. Alteraciones genéticas que afectan la estructura, el número o la especificidad de las porinas pueden modificar la permeabilidad de la membrana externa y dar lugar a fenómenos de resistencia antibiótica, al reducir aún más la entrada de fármacos hacia el interior celular.
Además de las porinas, la membrana externa contiene proteínas con funciones estructurales que contribuyen a la estabilidad mecánica de la envoltura celular. Otras proteínas actúan como receptores específicos para bacteriófagos, lo que condiciona la susceptibilidad de la bacteria a infecciones virales. También se encuentran presentes componentes de sistemas de transporte y de secreción, que permiten el movimiento controlado de sustancias hacia el interior o el exterior de la célula, así como la exportación de factores de virulencia durante la infección.
La membrana externa no es una estructura aislada, sino que se encuentra firmemente integrada con el resto de la envoltura celular. Está conectada con la membrana citoplasmática a través de sitios de adhesión especializados, que establecen una continuidad funcional entre ambas membranas. Estos sitios desempeñan un papel esencial en el transporte y ensamblaje de componentes recién sintetizados de la membrana externa, facilitando su entrega desde el citoplasma hasta su ubicación final.
Asimismo, la membrana externa está anclada al peptidoglucano mediante una lipoproteína específica. Esta lipoproteína se encuentra unida de forma covalente al peptidoglucano y, al mismo tiempo, insertada en la membrana externa. Esta doble unión asegura la cohesión estructural de la envoltura celular, integrando la capa rígida de peptidoglucano con la membrana externa y evitando su separación bajo condiciones de estrés mecánico u osmótico.
La estabilidad de la membrana externa depende también de interacciones químicas entre sus componentes. Los lipopolisacáridos de la cara externa de la membrana se encuentran unidos entre sí mediante puentes formados por cationes divalentes, principalmente magnesio y calcio, que enlazan los grupos fosfato de las moléculas de lipopolisacárido. A estas interacciones iónicas se suman interacciones hidrofóbicas entre los lipopolisacáridos y las proteínas de la membrana, generando una estructura rígida, compacta y altamente resistente. Esta organización confiere a la membrana externa una notable fortaleza física y una baja permeabilidad a numerosas sustancias potencialmente dañinas.
Sin embargo, esta estabilidad puede verse comprometida bajo ciertas condiciones. Algunos antibióticos, como las polimixinas, actúan directamente sobre la membrana externa, alterando sus interacciones estructurales. De igual modo, la eliminación de los cationes divalentes mediante agentes quelantes, como el ácido etilendiaminotetraacético o ciertos antibióticos, debilita los enlaces entre los lipopolisacáridos. La disrupción de estas interacciones provoca un aumento de la permeabilidad de la membrana externa, permitiendo la entrada de moléculas grandes o hidrofóbicas que normalmente serían excluidas.
Cuando la membrana externa se ve alterada, la bacteria pierde una de sus principales defensas. En estas condiciones, enzimas como la lisozima pueden acceder al peptidoglucano subyacente y degradarlo parcialmente. Este proceso da lugar a la formación de esferoplastos, células bacterianas que conservan parte de su envoltura, pero que, al igual que los protoplastos derivados de bacterias Gram positivas, son extremadamente sensibles a los cambios osmóticos. La pérdida de la integridad de la membrana externa y del soporte del peptidoglucano compromete de manera crítica la viabilidad bacteriana.

Fuente y lecturas recomendadas:
- Madigan, M. T., Martinko, J. M., Bender, K. S., Buckley, D. H., & Stahl, D. A. (2018). Brock biology of microorganisms (15th ed.). Pearson.
- Murray, P. R., Rosenthal, K. S., & Pfaller, M. A. (2025). Medical microbiology (10th ed.). Elsevier.
- Carroll, K. C., & Pfaller, M. A. (2023). Manual of clinical microbiology (13th ed.). American Society for Microbiology Press.
- Riedel, S., Hobden, J. A., Miller, S., Morse, S. A., Mietzner, T. A., Detrick, B., Mitchell, T. G., Sakanari, J. A., Hotez, P., & Mejía, R. (2020). Microbiología médica (28ª ed.). McGraw-Hill Interamericana Editores.
Aprende administración paso a paso

ADMINISTRACION DESDE CERO