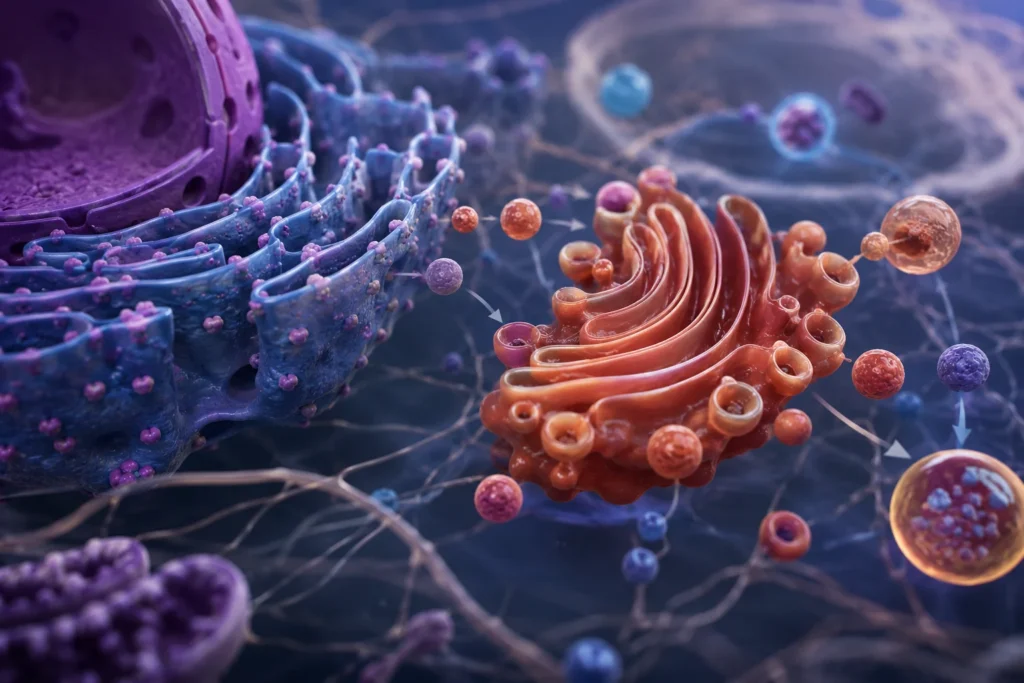
El retículo endoplásmico y el aparato de Golgi constituyen un sistema integrado de membranas intracelulares responsable de la biosíntesis, modificación, clasificación, transporte y distribución de una enorme variedad de moléculas esenciales para la supervivencia celular. En las células altamente secretoras, como las células acinares pancreáticas, los plasmocitos, los hepatocitos y numerosas células glandulares, estos orgánulos ocupan una proporción considerable del volumen citoplasmático debido a la intensa actividad metabólica necesaria para producir proteínas, lípidos, glucoproteínas, proteoglucanos y otros componentes celulares. Esta expansión estructural refleja directamente la elevada demanda de síntesis y tráfico intracelular de macromoléculas.
Las membranas que forman tanto el retículo endoplásmico como el aparato de Golgi están constituidas por bicapas lipídicas semejantes a la membrana plasmática. Estas bicapas contienen proteínas integrales y periféricas especializadas que funcionan como enzimas, receptores, transportadores y componentes estructurales. La organización de estas membranas permite crear compartimentos intracelulares con ambientes químicos específicos, favoreciendo la realización simultánea de múltiples reacciones metabólicas incompatibles entre sí si ocurrieran en el citosol.
Organización estructural y funcional del retículo endoplásmico
El retículo endoplásmico constituye la red membranosa más extensa de la célula eucariota. Se encuentra conectado físicamente con la envoltura nuclear y se distribuye a través de gran parte del citoplasma formando cisternas, túbulos y vesículas interconectadas.
Desde el punto de vista funcional y morfológico se divide en dos regiones principales: el retículo endoplásmico rugoso y el retículo endoplásmico liso.
La diferencia fundamental entre ambas regiones radica en la presencia o ausencia de ribosomas adheridos a la superficie citoplasmática de sus membranas. Esta característica determina sus funciones fisiológicas especializadas.
Síntesis de proteínas en el retículo endoplásmico rugoso
El retículo endoplásmico rugoso constituye el principal centro de síntesis de proteínas destinadas a la secreción extracelular, a la membrana plasmática y a los orgánulos del sistema endomembranoso.
Los ribosomas unidos a su superficie traducen moléculas de ARN mensajero mediante el proceso de síntesis proteica. Cuando una proteína posee una secuencia señal específica, el complejo ribosoma-ARN mensajero es dirigido hacia la membrana del retículo endoplásmico mediante la acción de la partícula de reconocimiento de señal. Esta interacción permite que la proteína naciente atraviese un canal denominado translocón y penetre simultáneamente en la luz del retículo mientras continúa su síntesis.
Este mecanismo recibe el nombre de translocación cotraduccional y constituye uno de los procesos fundamentales para la organización interna de las células eucariotas. Gracias a este sistema, las proteínas pueden ingresar directamente en el compartimento donde comenzarán su plegamiento y procesamiento inicial.
Una vez dentro de la luz del retículo endoplásmico, las proteínas recién sintetizadas experimentan modificaciones esenciales para alcanzar su conformación funcional. Entre estas modificaciones destacan la formación de puentes disulfuro, el plegamiento tridimensional asistido por chaperonas moleculares y la glucosilación inicial mediante la adición de oligosacáridos específicos.
Las proteínas mal plegadas son retenidas por mecanismos de control de calidad altamente especializados. Si no logran adquirir una conformación correcta, son transportadas nuevamente hacia el citosol para su degradación mediante el sistema ubiquitina-proteasoma. Este proceso evita la acumulación de proteínas defectuosas potencialmente tóxicas para la célula.
La relevancia fisiológica de este sistema es enorme. La mayoría de las hormonas proteicas, enzimas digestivas, anticuerpos, factores de coagulación, receptores de membrana y proteínas de transporte dependen de la maquinaria biosintética del retículo endoplásmico rugoso para su correcta producción.
Plegamiento proteico y control de calidad
El ambiente interno del retículo endoplásmico está especialmente adaptado para favorecer el plegamiento proteico. En su interior existe una elevada concentración de proteínas chaperonas, entre las que destacan BiP, calnexina y calreticulina.
Estas moléculas reconocen regiones hidrofóbicas expuestas en proteínas recién sintetizadas y previenen agregaciones incorrectas. Además, colaboran activamente en la adquisición de la estructura tridimensional definitiva.
Cuando la capacidad de plegamiento es superada por un exceso de proteínas defectuosas, se activa una respuesta adaptativa denominada respuesta a proteínas mal plegadas. Este mecanismo aumenta la producción de chaperonas, reduce temporalmente la síntesis proteica y estimula la eliminación de proteínas anómalas.
La activación prolongada de esta respuesta se ha asociado con enfermedades neurodegenerativas, diabetes mellitus, enfermedades hepáticas y diversos tipos de cáncer.
Síntesis de lípidos en el retículo endoplásmico liso
El retículo endoplásmico liso constituye el principal sitio celular de biosíntesis lipídica. Sus membranas contienen numerosas enzimas especializadas capaces de sintetizar fosfolípidos, colesterol, ceramidas, triglicéridos y otros componentes fundamentales de las membranas biológicas.
Los fosfolípidos sintetizados son incorporados inicialmente a la monocapa citoplasmática de la membrana del retículo. Posteriormente, proteínas denominadas flipasas, flopasas y escramblasas redistribuyen estas moléculas entre ambas monocapas para mantener la simetría y funcionalidad de la bicapa lipídica.
La síntesis continua de fosfolípidos provoca el crecimiento de las membranas reticulares. Parte de este material membranoso es transferido mediante vesículas hacia otros compartimentos celulares, contribuyendo a la renovación permanente de membranas intracelulares y de la membrana plasmática.
El colesterol sintetizado en el retículo endoplásmico desempeña funciones estructurales fundamentales. Modula la fluidez de las membranas, participa en la formación de microdominios lipídicos y constituye el precursor de hormonas esteroideas, ácidos biliares y vitamina D.
En células endocrinas especializadas, el retículo endoplásmico liso alcanza un desarrollo extraordinario debido a la intensa producción de hormonas esteroideas.
Participación en el metabolismo del glucógeno
El retículo endoplásmico liso contiene enzimas involucradas en el metabolismo de los carbohidratos, particularmente en los hepatocitos.
Una de las más importantes es la glucosa-6-fosfatasa, localizada en la membrana del retículo endoplásmico. Esta enzima cataliza la conversión de glucosa-6-fosfato en glucosa libre, permitiendo que la glucosa producida durante la glucogenólisis y la gluconeogénesis pueda ser liberada al torrente sanguíneo.
Este mecanismo es esencial para mantener la homeostasis de la glucosa durante periodos de ayuno y constituye una función metabólica exclusiva de determinados tejidos.
Detoxificación de sustancias potencialmente dañinas
Otra función crucial del retículo endoplásmico liso consiste en la biotransformación de compuestos tóxicos.
Las membranas reticulares contienen enzimas pertenecientes a la superfamilia del citocromo P450, capaces de oxidar una amplia variedad de sustancias exógenas y endógenas.
Estas enzimas convierten moléculas liposolubles en compuestos más hidrosolubles que pueden eliminarse con mayor facilidad por vía renal o biliar.
Los procesos de detoxificación incluyen oxidación, reducción, hidrólisis y reacciones de conjugación con glucurónido, sulfato, glutatión u otras moléculas.
La exposición crónica a determinados fármacos puede inducir una expansión significativa del retículo endoplásmico liso, aumentando la capacidad metabólica de la célula para procesar dichas sustancias.
Transporte vesicular desde el retículo endoplásmico hacia el aparato de Golgi
Las proteínas y lípidos sintetizados en el retículo endoplásmico deben ser transportados hacia otros compartimentos celulares para completar su procesamiento.
Este transporte se realiza mediante vesículas recubiertas por proteínas especializadas, principalmente complejos COPII.
Las vesículas brotan desde regiones específicas del retículo endoplásmico conocidas como sitios de salida. Posteriormente se desplazan hacia el compartimento intermedio retículo endoplásmico-Golgi y finalmente alcanzan la cara cis del aparato de Golgi.
La especificidad de este tráfico depende de proteínas Rab, proteínas de anclaje y complejos SNARE, que garantizan la fusión correcta entre membranas.
Este sistema asegura que cada molécula alcance exactamente el compartimento donde debe continuar su procesamiento.
Organización estructural del aparato de Golgi
El aparato de Golgi está constituido por una serie de cisternas membranosas aplanadas organizadas en pilas.
Posee una polaridad estructural claramente definida. La cara cis recibe materiales procedentes del retículo endoplásmico, mientras que la cara trans actúa como centro de clasificación y distribución.
Entre ambos extremos existen cisternas intermedias donde se llevan a cabo numerosas modificaciones bioquímicas.
Esta organización secuencial permite que las moléculas experimenten cambios progresivos a medida que avanzan a través del orgánulo.
Modificaciones de proteínas en el aparato de Golgi
Una de las funciones más importantes del aparato de Golgi consiste en modificar proteínas sintetizadas previamente en el retículo endoplásmico.
Entre las modificaciones más relevantes se encuentran la remodelación de cadenas de oligosacáridos, la glucosilación adicional, la fosforilación, la sulfatación y diversos procesos de proteólisis limitada.
Estas modificaciones alteran la estabilidad, actividad biológica, localización intracelular y capacidad de interacción de las proteínas.
La glucosilación es particularmente importante porque participa en el reconocimiento celular, la adhesión intercelular, la respuesta inmunitaria y el tráfico intracelular.
Muchas proteínas no alcanzan su funcionalidad definitiva hasta completar estas modificaciones en el aparato de Golgi.
Síntesis de polisacáridos y proteoglucanos
Además de modificar proteínas, el aparato de Golgi sintetiza numerosos carbohidratos complejos.
Entre los productos más importantes se encuentran los glucosaminoglucanos y proteoglucanos de la matriz extracelular.
El ácido hialurónico, el sulfato de condroitina, el sulfato de dermatán, el sulfato de heparán y otras macromoléculas relacionadas desempeñan funciones fundamentales en la arquitectura tisular.
Estas moléculas forman geles altamente hidratados que proporcionan resistencia mecánica, elasticidad y capacidad de difusión de nutrientes.
En el cartílago, los proteoglucanos permiten soportar fuerzas compresivas intensas. En la matriz extracelular general regulan la migración celular, la proliferación, la diferenciación y numerosos procesos de señalización.
Durante el desarrollo embrionario, la reparación tisular y la cicatrización, estas moléculas participan activamente en la organización espacial de las células y en la formación de nuevos tejidos.
Compactación y clasificación de productos secretorios
A medida que las moléculas avanzan a través de las cisternas del aparato de Golgi, sufren una progresiva concentración y empaquetamiento.
Las proteínas son clasificadas según señales moleculares específicas que determinan su destino final.
Algunas serán secretadas al exterior celular mediante exocitosis constitutiva o regulada. Otras serán incorporadas a membranas celulares. Un tercer grupo será enviado a lisosomas u otros orgánulos intracelulares.
Esta clasificación representa uno de los mecanismos más sofisticados de organización celular.
Formación de vesículas secretoras
Las células secretoras producen grandes cantidades de vesículas especializadas destinadas a liberar sustancias al espacio extracelular.
Estas vesículas se originan principalmente en la red trans-Golgi, donde las proteínas son concentradas y empaquetadas.
Las vesículas secretoras almacenan hormonas, neurotransmisores, enzimas digestivas, mucinas, anticuerpos y numerosas moléculas biológicamente activas.
Cuando reciben señales adecuadas, migran hacia la membrana plasmática y se preparan para fusionarse con ella.
Exocitosis y papel del calcio
La exocitosis constituye el mecanismo mediante el cual las células liberan materiales al medio extracelular.
En muchas células secretoras, el aumento de la concentración intracelular de calcio actúa como señal desencadenante.
El calcio interactúa con proteínas sensoras presentes en las membranas vesiculares, especialmente sinaptotagminas. Esta interacción activa complejos SNARE que aproximan las membranas hasta provocar su fusión.
Como resultado, el contenido vesicular es liberado al exterior mientras la membrana de la vesícula se incorpora temporalmente a la membrana plasmática.
Este mecanismo es esencial para la neurotransmisión, la secreción hormonal, la respuesta inmunitaria y numerosos procesos fisiológicos.
Formación de lisosomas
El aparato de Golgi también participa en la generación de lisosomas.
Las hidrolasas lisosomales sintetizadas en el retículo endoplásmico reciben en el aparato de Golgi una señal molecular específica denominada manosa-6-fosfato.
Esta etiqueta permite que las enzimas sean reconocidas por receptores especializados y dirigidas hacia compartimentos endosómicos que finalmente maduran hasta convertirse en lisosomas.
Los lisosomas contienen numerosas enzimas hidrolíticas capaces de degradar proteínas, lípidos, carbohidratos y ácidos nucleicos.
Su función es esencial para el reciclaje de componentes celulares y la eliminación de materiales dañados.
Renovación y expansión de membranas celulares
Las vesículas originadas en el aparato de Golgi no solo participan en la secreción.
Una proporción importante se fusiona con membranas intracelulares y con la membrana plasmática, contribuyendo a su mantenimiento y renovación.
Este proceso resulta indispensable porque las membranas celulares sufren una constante pérdida de material debido a fenómenos de endocitosis, fagocitosis, pinocitosis y recambio molecular.
El aporte continuo de nuevos lípidos y proteínas permite conservar la integridad estructural de todos los compartimentos celulares.
Importancia fisiológica del sistema retículo endoplásmico-Golgi
El sistema integrado formado por el retículo endoplásmico y el aparato de Golgi representa uno de los centros metabólicos más activos de la célula eucariota.
Su actividad permite la producción coordinada de proteínas, lípidos, carbohidratos complejos, membranas celulares y orgánulos especializados.
Además, controla rigurosamente la calidad de las macromoléculas sintetizadas, regula su distribución intracelular y asegura que cada producto alcance su destino funcional correcto.
La alteración de cualquiera de estos procesos puede producir enfermedades metabólicas, neurodegenerativas, inmunológicas, endocrinas y neoplásicas, lo que evidencia la importancia crítica de estas estructuras para la homeostasis celular y la fisiología de los organismos multicelulares.


Fuente y lecturas recomendadas:
- Alberts, B., Johnson, A., Lewis, J., Morgan, D., Raff, M., Roberts, K., & Walter, P. (2022). Molecular biology of the cell (7th ed.). W. W. Norton & Company.
- Braakman, I., & Hebert, D. N. (2013). Protein folding in the endoplasmic reticulum. Cold Spring Harbor Perspectives in Biology, 5(5), a013201. https://doi.org/10.1101/cshperspect.a013201
- Ghaemmaghami, S., & Kaufman, R. J. (2004). Endoplasmic reticulum stress and the unfolded protein response: Balancing cell life and death. Current Opinion in Cell Biology, 16(4), 451–457. https://doi.org/10.1016/j.ceb.2004.06.007
- Hetz, C., Zhang, K., & Kaufman, R. J. (2020). Mechanisms, regulation and functions of the unfolded protein response. Nature Reviews Molecular Cell Biology, 21(8), 421–438. https://doi.org/10.1038/s41580-020-0250-z
- Lippincott-Schwartz, J., Roberts, T. H., & Hirschberg, K. (2000). Secretory protein trafficking and organelle dynamics in living cells. Annual Review of Cell and Developmental Biology, 16, 557–589. https://doi.org/10.1146/annurev.cellbio.16.1.557
- Lodish, H., Berk, A., Kaiser, C. A., Krieger, M., Bretscher, A., Ploegh, H., Amon, A., & Scott, M. P. (2021). Molecular cell biology (9th ed.). W. H. Freeman.
- Mellman, I., & Warren, G. (2000). The road taken: Past and future foundations of membrane traffic. Cell, 100(1), 99–112. https://doi.org/10.1016/S0092-8674(00)81686-2
- Molinari, M. (2007). N-glycan structure dictates extension of protein folding or onset of disposal. Nature Chemical Biology, 3(6), 313–320. https://doi.org/10.1038/nchembio0607-313
- Rothman, J. E., & Wieland, F. T. (1996). Protein sorting by transport vesicles. Science, 272(5259), 227–234. https://doi.org/10.1126/science.272.5259.227
- Stanley, P. (2011). Golgi glycosylation. Cold Spring Harbor Perspectives in Biology, 3(4), a005199. https://doi.org/10.1101/cshperspect.a005199
- Voeltz, G. K., Rolls, M. M., & Rapoport, T. A. (2002). Structural organization of the endoplasmic reticulum. EMBO Reports, 3(10), 944–950. https://doi.org/10.1093/embo-reports/kvf202
- Walter, P., & Ron, D. (2011). The unfolded protein response: From stress pathway to homeostatic regulation. Science, 334(6059), 1081–1086. https://doi.org/10.1126/science.1209038
- Wang, Y., & Seemann, J. (2011). Golgi biogenesis. Cold Spring Harbor Perspectives in Biology, 3(10), a005330. https://doi.org/10.1101/cshperspect.a005330
- Weerapana, E., & Imperiali, B. (2006). Asparagine-linked protein glycosylation: From eukaryotic to prokaryotic systems. Glycobiology, 16(6), 91R–101R. https://doi.org/10.1093/glycob/cwj099
- Zerial, M., & McBride, H. (2001). Rab proteins as membrane organizers. Nature Reviews Molecular Cell Biology, 2(2), 107–117. https://doi.org/10.1038/35052055
Aprende administración paso a paso

ADMINISTRACION DESDE CERO