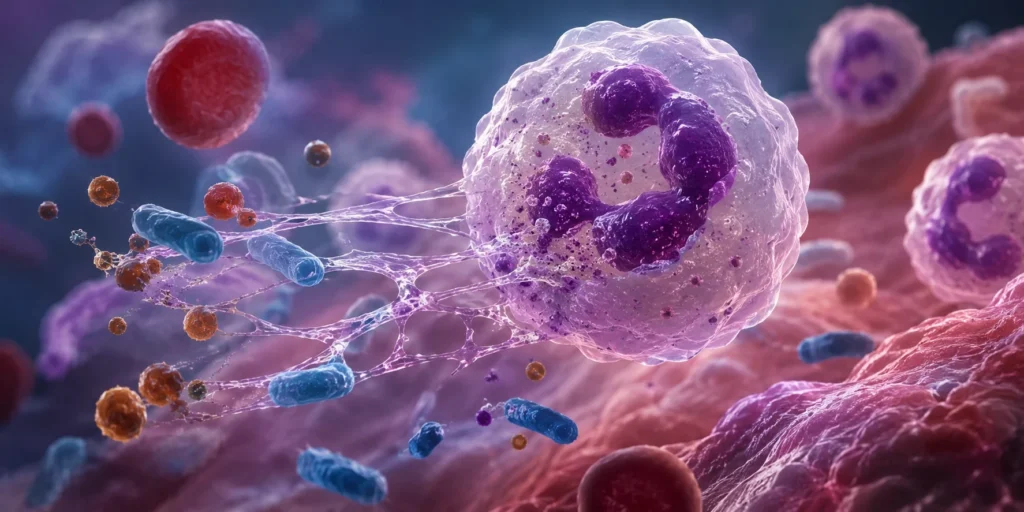
El neutrófilo es el leucocito más abundante de la sangre periférica humana y constituye el principal componente celular de la respuesta inmunitaria innata durante las fases iniciales de la inflamación aguda. Pertenece al grupo de los leucocitos polimorfonucleares debido a la morfología característica de su núcleo, el cual presenta entre 2 y 5 lóbulos unidos por delgados puentes de cromatina. Esta configuración nuclear no representa únicamente una característica morfológica, sino una adaptación funcional que incrementa la flexibilidad celular y facilita el desplazamiento del neutrófilo a través de espacios intercelulares extremadamente estrechos durante el proceso de extravasación hacia los tejidos lesionados. La elevada deformabilidad del núcleo, junto con un citoesqueleto dinámico constituido por filamentos de actina, microtúbulos y proteínas motoras, permite que estas células atraviesen rápidamente el endotelio vascular y migren hacia el sitio donde se desarrolla el daño tisular o la invasión microbiana. Esta capacidad migratoria constituye una de las razones fundamentales por las cuales los neutrófilos son las primeras células inflamatorias en acumularse en los focos de inflamación aguda.
La rápida participación de los neutrófilos obedece a diversos factores fisiológicos. En primer lugar, existe un gran reservorio de neutrófilos maduros almacenados en la médula ósea, cuya magnitud supera ampliamente el número de neutrófilos circulantes. En respuesta a estímulos inflamatorios, citocinas y factores estimulantes de colonias favorecen la liberación masiva de estas células hacia la circulación sanguínea, fenómeno denominado desmarginalización y movilización medular. En segundo lugar, los neutrófilos expresan de manera constitutiva un amplio repertorio de receptores para quimiocinas, moléculas derivadas del complemento, productos bacterianos y señales emitidas por células lesionadas, lo que les permite detectar rápidamente gradientes quimiotácticos y dirigirse con precisión hacia el sitio de inflamación. Finalmente, las células endoteliales activadas expresan moléculas de adhesión que interactúan específicamente con receptores presentes en la superficie del neutrófilo, facilitando su adhesión, rodamiento, activación y migración transendotelial.
Durante las primeras horas posteriores a una lesión tisular o al establecimiento de una infección, los tejidos afectados liberan numerosas moléculas conocidas como patrones moleculares asociados con daño y patrones moleculares asociados con patógenos. Estas señales son reconocidas por receptores de reconocimiento de patrones presentes en macrófagos residentes, células dendríticas, mastocitos y células epiteliales. Como consecuencia, estas células sintetizan mediadores inflamatorios que incluyen factor de necrosis tumoral, interleucina 1, interleucina 6, interleucina 8, leucotrieno B4, prostaglandinas, histamina y quimiocinas específicas. Dichos mediadores producen vasodilatación, incremento de la permeabilidad vascular y activación del endotelio, preparando el lecho vascular para el reclutamiento masivo de neutrófilos.
El proceso mediante el cual el neutrófilo abandona la circulación recibe el nombre de cascada de adhesión leucocitaria. Inicialmente ocurre el rodamiento sobre la superficie endotelial, mediado principalmente por selectinas. Posteriormente, las quimiocinas inmovilizadas sobre la superficie endotelial activan receptores específicos del neutrófilo, induciendo un cambio conformacional en las integrinas de alta afinidad. Estas integrinas permiten una adhesión firme mediante su interacción con moléculas de adhesión intercelular expresadas por el endotelio activado. Finalmente, el neutrófilo atraviesa las uniones intercelulares del endotelio mediante diapédesis y continúa desplazándose por el tejido siguiendo gradientes crecientes de factores quimiotácticos.
La migración dirigida del neutrófilo depende principalmente de la quimiotaxis, proceso mediante el cual la célula detecta diferencias extremadamente pequeñas en la concentración de moléculas señalizadoras distribuidas en el tejido inflamado. Entre los quimioatrayentes más potentes destacan el péptido bacteriano N-formilmetionil-leucil-fenilalanina, el fragmento C5a del complemento, el leucotrieno B4, la quimiocina CXCL8 y diversas proteínas liberadas por células necróticas. La unión de estos ligandos a receptores acoplados con proteínas G activa múltiples vías intracelulares que reorganizan el citoesqueleto y establecen una polaridad funcional caracterizada por la formación de un borde de avance rico en actina y un extremo posterior contráctil, permitiendo una migración altamente eficiente.
Una vez que alcanza el sitio inflamatorio, el neutrófilo se convierte en uno de los principales efectores de la respuesta inmunitaria. Su función primordial consiste en eliminar microorganismos mediante fagocitosis, desgranulación y producción de especies reactivas de oxígeno. La fagocitosis inicia con el reconocimiento del microorganismo a través de receptores para inmunoglobulina G, receptores para componentes del complemento y receptores de reconocimiento de patrones. La internalización del microorganismo conduce a la formación del fagosoma, el cual posteriormente se fusiona con los gránulos citoplasmáticos para formar el fagolisosoma, donde ocurre la destrucción del agente infeccioso.
El citoplasma del neutrófilo contiene varios tipos de gránulos especializados. Los gránulos primarios o azurófilos contienen mieloperoxidasa, elastasa, catepsina G, proteinasa 3, defensinas y diversas hidrolasas ácidas. Los gránulos secundarios contienen lactoferrina, lisozima y componentes de la nicotinamida adenina dinucleótido fosfato oxidasa. Los gránulos terciarios contienen metaloproteinasas que facilitan la degradación de la matriz extracelular y favorecen la migración celular. La liberación regulada de estas proteínas permite destruir microorganismos, aunque también puede producir lesión tisular cuando la respuesta inflamatoria resulta excesiva o prolongada.
La producción de especies reactivas de oxígeno representa otro mecanismo fundamental de defensa. La activación del complejo nicotinamida adenina dinucleótido fosfato oxidasa genera anión superóxido, que posteriormente origina peróxido de hidrógeno. La mieloperoxidasa utiliza este peróxido para producir ácido hipocloroso, uno de los agentes microbicidas más potentes sintetizados por las células humanas. Estas especies reactivas oxidan proteínas, lípidos y ácidos nucleicos de los microorganismos, contribuyendo de manera decisiva a su destrucción.
Además de la fagocitosis, los neutrófilos pueden liberar trampas extracelulares compuestas por cromatina descondensada asociada con proteínas antimicrobianas. Estas estructuras inmovilizan bacterias, hongos y algunos virus, concentrando localmente enzimas microbicidas que limitan la diseminación del agente infeccioso. Sin embargo, cuando este mecanismo ocurre de forma exagerada puede favorecer trombosis, lesión endotelial, enfermedades autoinmunes y daño tisular persistente.
Aunque históricamente se consideró que los neutrófilos eran células exclusivamente efectoras y de vida muy corta, actualmente se reconoce que poseen una notable capacidad inmunorreguladora. Estas células producen numerosas citocinas, quimiocinas, factores angiogénicos, mediadores lipídicos y vesículas extracelulares que modulan la actividad de macrófagos, células dendríticas, linfocitos T, linfocitos B y células endoteliales. De esta manera, el neutrófilo no solamente elimina microorganismos, sino que también coordina la intensidad y duración de la respuesta inflamatoria.
En condiciones fisiológicas, los neutrófilos circulantes presentan una semivida aproximada de 6 a 8 horas en la sangre periférica. Esta corta permanencia constituye un mecanismo homeostático que limita el riesgo de daño tisular derivado de la liberación espontánea de enzimas proteolíticas y especies reactivas de oxígeno. Después de este periodo, los neutrófilos experimentan apoptosis espontánea, un proceso programado que evita la liberación descontrolada de su contenido citotóxico. Posteriormente son eliminados por macrófagos residentes en médula ósea, hígado, bazo y otros tejidos mediante eferocitosis, mecanismo que contribuye a la resolución de la inflamación.
No obstante, la longevidad del neutrófilo aumenta significativamente durante los procesos inflamatorios. Diversas citocinas, factores de crecimiento y productos microbianos activan vías intracelulares que retrasan la apoptosis. Entre los principales mediadores destacan el factor estimulante de colonias de granulocitos, el factor estimulante de colonias de granulocitos y macrófagos, el factor de necrosis tumoral, la interleucina 1β, la interleucina 6, el interferón γ, diversas quimiocinas y componentes bacterianos como el lipopolisacárido. Estas señales incrementan la expresión de proteínas antiapoptóticas de la familia BCL-2, especialmente MCL-1 y A1, mientras inhiben la activación de caspasas responsables de ejecutar el programa apoptótico.
El aumento de la supervivencia del neutrófilo posee un claro significado biológico. Durante una infección grave, la permanencia prolongada de estas células permite mantener una elevada capacidad microbicida mientras persiste el estímulo inflamatorio. Asimismo, la supervivencia prolongada favorece la producción sostenida de mediadores inmunitarios, incrementa la capacidad fagocítica y optimiza la coordinación con otros componentes de la respuesta inmunitaria. Sin embargo, cuando este mecanismo persiste más allá de lo necesario puede contribuir al desarrollo de inflamación crónica, lesión tisular progresiva y enfermedades inmunomediadas.
En enfermedades inflamatorias como artritis reumatoide, enfermedad pulmonar obstructiva crónica, síndrome de dificultad respiratoria aguda, vasculitis asociadas con anticuerpos anticitoplasma de neutrófilos, psoriasis y diversas enfermedades cardiovasculares, la prolongación anormal de la supervivencia de los neutrófilos favorece la acumulación persistente de enzimas proteolíticas, radicales libres y trampas extracelulares, amplificando el daño tisular y perpetuando el proceso inflamatorio. Por el contrario, en estados de neutropenia o alteraciones funcionales del neutrófilo, la disminución del número o de la capacidad microbicida de estas células incrementa considerablemente la susceptibilidad a infecciones bacterianas y fúngicas invasoras.
La resolución adecuada de la inflamación depende finalmente de la eliminación ordenada de los neutrófilos mediante apoptosis seguida de eferocitosis. Este proceso induce un cambio funcional en los macrófagos hacia un fenotipo reparador, favorece la producción de mediadores proresolutivos, disminuye la síntesis de citocinas proinflamatorias y promueve la reparación del tejido lesionado. Por ello, la duración de la vida del neutrófilo constituye un punto de regulación esencial que determina el equilibrio entre una respuesta inmunitaria eficaz frente a los microorganismos y la prevención del daño inflamatorio excesivo.


Fuente y lecturas recomendadas:
- Borregaard, N. (2010). Neutrophils, from marrow to microbes. Immunity, 33(5), 657-670. https://doi.org/10.1016/j.immuni.2010.11.011
- Brinkmann, V., Reichard, U., Goosmann, C., Fauler, B., Uhlemann, Y., Weiss, D. S., Weinrauch, Y., & Zychlinsky, A. (2004). Neutrophil extracellular traps kill bacteria. Science, 303(5663), 1532-1535. https://doi.org/10.1126/science.1092385
- Burn, G. L., Foti, A., Marsman, G., Patel, D. F., & Zychlinsky, A. (2021). The neutrophil. Immunity, 54(7), 1377-1391. https://doi.org/10.1016/j.immuni.2021.06.006
- Kolaczkowska, E., & Kubes, P. (2013). Neutrophil recruitment and function in health and inflammation. Nature Reviews Immunology, 13(3), 159-175. https://doi.org/10.1038/nri3399
- Lawrence, S. M., Corriden, R., & Nizet, V. (2018). The ontogeny of a neutrophil: Mechanisms of granulopoiesis and homeostasis. Microbiology and Molecular Biology Reviews, 82(1), e00057-17. https://doi.org/10.1128/MMBR.00057-17
- Nathan, C. (2006). Neutrophils and immunity: Challenges and opportunities. Nature Reviews Immunology, 6(3), 173-182. https://doi.org/10.1038/nri1785
- Nauseef, W. M. (2007). How human neutrophils kill and degrade microbes: An integrated view. Immunological Reviews, 219(1), 88-102. https://doi.org/10.1111/j.1600-065X.2007.00550.x
- Pillay, J., den Braber, I., Vrisekoop, N., Kwast, L. M., de Boer, R. J., Borghans, J. A. M., Tesselaar, K., & Koenderman, L. (2010). In vivo labeling with ²H₂O reveals a human neutrophil lifespan of 5.4 days. Blood, 116(4), 625-627. https://doi.org/10.1182/blood-2010-01-259028
- Summers, C., Rankin, S. M., Condliffe, A. M., Singh, N., Peters, A. M., & Chilvers, E. R. (2010). Neutrophil kinetics in health and disease. Trends in Immunology, 31(8), 318-324. https://doi.org/10.1016/j.it.2010.05.006
- Wright, H. L., Moots, R. J., Bucknall, R. C., & Edwards, S. W. (2010). Neutrophil function in inflammation and inflammatory diseases. Rheumatology, 49(9), 1618-1631. https://doi.org/10.1093/rheumatology/keq045
Aprende administración paso a paso

ADMINISTRACION DESDE CERO