
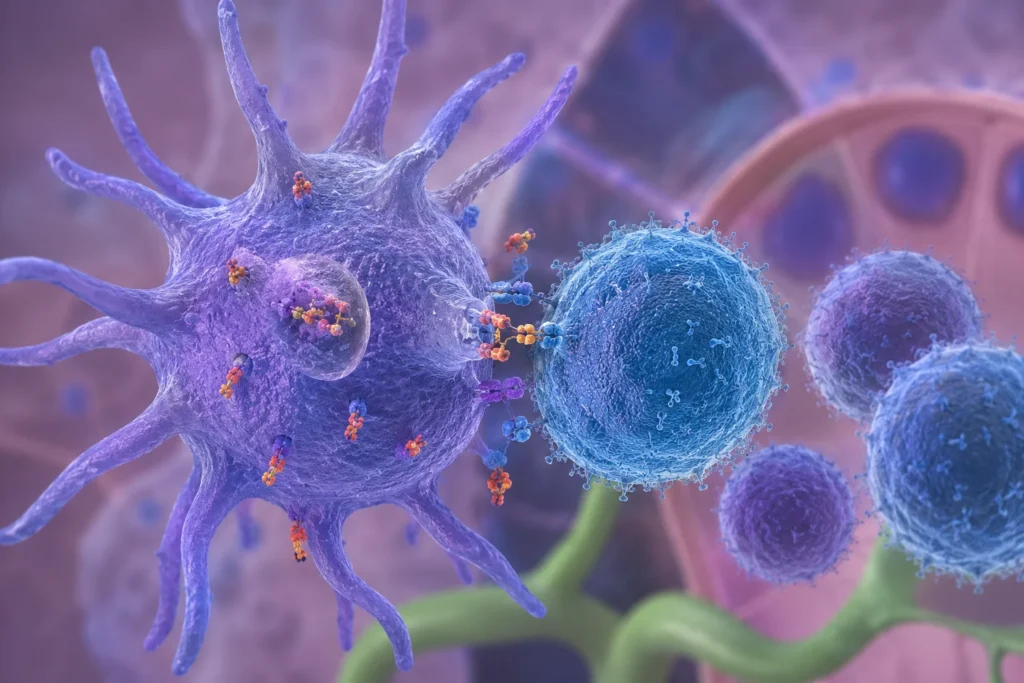
Las células dendríticas constituyen el principal vínculo funcional entre la inmunidad innata y la inmunidad adaptativa porque poseen la capacidad única de detectar microorganismos invasores o moléculas extrañas, procesar sus antígenos y activar linfocitos T vírgenes, iniciando así una respuesta inmunitaria específica y de memoria. Ninguna otra célula del sistema inmunitario reúne simultáneamente una capacidad tan elevada para reconocer patrones moleculares asociados con patógenos, captar antígenos, procesarlos, migrar hacia órganos linfoides secundarios y proporcionar todas las señales necesarias para la activación completa de los linfocitos T. Esta combinación de funciones convierte a las células dendríticas en las células presentadoras de antígeno profesionales más eficientes del organismo.
Las células dendríticas se originan en la médula ósea a partir de progenitores hematopoyéticos y se distribuyen ampliamente en prácticamente todos los tejidos del organismo. Son especialmente abundantes en aquellos sitios que representan las principales puertas de entrada para microorganismos, como la piel, las mucosas del aparato respiratorio, el aparato digestivo y el aparato genitourinario. Esta localización estratégica les permite actuar como centinelas inmunológicos, vigilando continuamente el ambiente extracelular en busca de señales que indiquen la presencia de microorganismos patógenos o daño tisular. Su estado fisiológico en los tejidos periféricos corresponde al de células dendríticas inmaduras, caracterizadas por una extraordinaria capacidad de captación de antígenos y una baja capacidad para estimular linfocitos T. Esta distribución anatómica y funcional permite que la respuesta inmunitaria se inicie inmediatamente después del ingreso de un agente infeccioso.
La primera función que establece el puente entre inmunidad innata y adaptativa consiste en el reconocimiento inicial del material extraño. Las células dendríticas expresan numerosos receptores de reconocimiento de patrones capaces de identificar estructuras moleculares altamente conservadas presentes en bacterias, virus, hongos y parásitos, conocidas como patrones moleculares asociados con patógenos. Asimismo, detectan moléculas liberadas por células dañadas o necróticas, denominadas patrones moleculares asociados con daño. Entre estos receptores destacan los receptores tipo Toll, los receptores tipo lectina C, los receptores tipo NOD, los receptores tipo RIG-I y diversos receptores captadores. La unión de estos receptores con sus ligandos activa complejas cascadas de señalización intracelular que inducen la maduración funcional de la célula dendrítica, la producción de citocinas inflamatorias y el incremento de la expresión de moléculas coestimuladoras indispensables para activar linfocitos T.
Una vez reconocido el agente extraño, la célula dendrítica incorpora el material antigénico mediante diversos mecanismos de endocitosis. Dependiendo de las características físicas y químicas del antígeno, puede utilizar fagocitosis para partículas grandes, macropinocitosis para captar grandes volúmenes de líquido extracelular o endocitosis mediada por receptores cuando existen ligandos específicos reconocidos por receptores de membrana. Durante esta etapa, las células dendríticas inmaduras presentan una actividad endocítica muy elevada, lo que les permite concentrar grandes cantidades de antígenos provenientes del ambiente donde residen.
Después de la internalización, el material antigénico es transportado hacia compartimentos intracelulares especializados donde ocurre su procesamiento. Las proteínas derivadas de microorganismos son degradadas progresivamente mediante enzimas proteolíticas presentes en endosomas y lisosomas, produciendo fragmentos peptídicos de longitud adecuada para ser presentados por las moléculas del complejo principal de histocompatibilidad. Este procesamiento constituye un requisito indispensable porque los receptores de los linfocitos T únicamente reconocen pequeños péptidos unidos a moléculas del complejo principal de histocompatibilidad y no proteínas intactas.
La vía de procesamiento depende principalmente del origen intracelular o extracelular del antígeno. Los antígenos provenientes del exterior de la célula, como bacterias extracelulares, toxinas o proteínas solubles, son procesados en compartimentos endolisosomales y posteriormente cargados sobre moléculas del complejo principal de histocompatibilidad de clase II. Estas moléculas se sintetizan inicialmente en el retículo endoplásmico asociadas con la cadena invariante, cuya función consiste en impedir la unión prematura de péptidos durante el transporte hacia los compartimentos endosomales. Una vez allí, la cadena invariante es degradada y sustituida por los péptidos antigénicos mediante la acción de moléculas especializadas que facilitan el intercambio peptídico. Finalmente, los complejos antígeno-complejo principal de histocompatibilidad clase II son transportados hacia la superficie celular.
Los antígenos sintetizados dentro de células infectadas, principalmente proteínas virales o proteínas derivadas de microorganismos intracelulares, siguen una vía diferente. Estas proteínas son degradadas por el proteasoma citoplasmático, cuyos productos peptídicos son transportados hacia el retículo endoplásmico mediante proteínas transportadoras asociadas con el procesamiento antigénico. Allí se unen a moléculas del complejo principal de histocompatibilidad de clase I, las cuales posteriormente migran hacia la superficie celular para ser reconocidas por linfocitos T citotóxicos.
Una característica extraordinaria de las células dendríticas consiste en la capacidad de realizar presentación cruzada. Este mecanismo permite que antígenos originalmente extracelulares sean presentados mediante moléculas del complejo principal de histocompatibilidad clase I, algo que normalmente ocurre únicamente con proteínas sintetizadas dentro de la célula. Gracias a este proceso, las células dendríticas pueden activar linfocitos T citotóxicos incluso cuando ellas mismas no están infectadas por el microorganismo. Este mecanismo resulta esencial para desarrollar inmunidad contra numerosos virus, algunos tumores y microorganismos intracelulares que no infectan directamente a las células dendríticas.
Mientras ocurre el procesamiento antigénico, la célula dendrítica experimenta un profundo proceso de maduración. Disminuye considerablemente su capacidad de captación de antígenos, pero incrementa de forma muy importante la expresión de moléculas del complejo principal de histocompatibilidad clase I y clase II, así como de moléculas coestimuladoras tales como CD80, CD86 y CD40. También aumenta la producción de citocinas inmunomoduladoras como interleucina doce, interferones tipo I, interleucina seis y otras moléculas que determinarán el tipo de respuesta inmunitaria adaptativa que se desarrollará posteriormente.
Simultáneamente, las células dendríticas modifican profundamente su patrón de expresión de receptores para quimiocinas. Durante el estado inmaduro predominan receptores que favorecen la permanencia en tejidos periféricos. Después de reconocer un patógeno, disminuye la expresión de dichos receptores y aumenta notablemente la expresión del receptor CCR7, cuya función consiste en responder a las quimiocinas producidas por vasos linfáticos y órganos linfoides secundarios. Como consecuencia, las células dendríticas abandonan el tejido donde captaron el antígeno y migran activamente a través de los vasos linfáticos aferentes hacia los ganglios linfáticos regionales. Cuando el antígeno circula por vía sanguínea, las células dendríticas también pueden dirigirse al bazo, donde cumplen funciones equivalentes.
Esta migración constituye uno de los aspectos más importantes que explican por qué las células dendríticas enlazan la inmunidad innata con la adaptativa. Los microorganismos suelen ingresar inicialmente en tejidos periféricos, mientras que la mayoría de los linfocitos T vírgenes permanecen circulando entre sangre y órganos linfoides secundarios. Por ello, es indispensable que exista una célula especializada capaz de transportar la información antigénica desde el sitio de infección hasta los lugares donde residen los linfocitos T. Sin este mecanismo de transporte, el encuentro entre ambas poblaciones celulares sería extremadamente ineficiente y la respuesta inmunitaria adaptativa difícilmente podría iniciarse.
Dentro de los ganglios linfáticos o del bazo, las células dendríticas maduras establecen contactos prolongados con miles de linfocitos T vírgenes mediante un proceso altamente organizado. Los linfocitos T exploran continuamente la superficie de múltiples células dendríticas utilizando sus receptores específicos para antígenos. Cuando un receptor reconoce un complejo formado por un péptido antigénico unido al complejo principal de histocompatibilidad apropiado, se inicia la formación de la sinapsis inmunológica, una estructura especializada que permite la comunicación estable entre ambas células.
La activación completa del linfocito T requiere tres señales complementarias. La primera corresponde al reconocimiento específico del complejo antígeno-complejo principal de histocompatibilidad por el receptor del linfocito T. Esta señal proporciona especificidad antigénica, pero por sí sola resulta insuficiente para inducir una respuesta inmunitaria efectiva.
La segunda señal corresponde a la coestimulación, proporcionada principalmente por la interacción entre las moléculas CD80 y CD86 presentes en la célula dendrítica con el receptor CD28 expresado en el linfocito T. La ausencia de esta segunda señal provoca un estado de anergia o tolerancia inmunológica, evitando respuestas contra antígenos inocuos o componentes propios del organismo. De esta manera, las células dendríticas desempeñan un papel fundamental tanto en la inducción de inmunidad como en el mantenimiento de la tolerancia inmunológica.
La tercera señal está constituida por las citocinas secretadas por la célula dendrítica durante la presentación antigénica. Estas moléculas determinan la diferenciación funcional del linfocito T activado. La producción predominante de interleucina doce favorece la diferenciación hacia linfocitos T cooperadores tipo 1, especializados en combatir microorganismos intracelulares. La presencia de otras citocinas favorece la generación de linfocitos T cooperadores tipo 2, tipo 17 o linfocitos T reguladores, cada uno adaptado para enfrentar diferentes clases de patógenos o mantener la homeostasis inmunológica.
Cuando el antígeno es presentado mediante moléculas del complejo principal de histocompatibilidad clase I, los linfocitos T CD8 vírgenes se activan y proliferan hasta diferenciarse en linfocitos T citotóxicos. Estas células adquieren la capacidad de reconocer y destruir específicamente células infectadas por virus, células parasitadas por microorganismos intracelulares y células tumorales mediante la liberación de perforinas, granzimas y otras moléculas citotóxicas. Gracias a este proceso, la respuesta inmunitaria adaptativa elimina células que albergan microorganismos inaccesibles para los anticuerpos.
Cuando la presentación ocurre mediante moléculas del complejo principal de histocompatibilidad clase II, los linfocitos T CD4 vírgenes se diferencian en linfocitos T cooperadores. Estas células coordinan prácticamente todos los componentes de la respuesta inmunitaria adaptativa mediante la secreción de citocinas que activan macrófagos, estimulan linfocitos B para producir anticuerpos, favorecen la expansión de linfocitos T citotóxicos y regulan la intensidad de la inflamación. De esta forma, una única célula dendrítica puede iniciar una respuesta inmunitaria compleja que involucra múltiples poblaciones celulares especializadas.
Las células dendríticas también participan activamente en la generación de memoria inmunológica. Durante la activación inicial de los linfocitos T, proporcionan señales que favorecen la formación de linfocitos T de memoria de larga duración. Estas células permanecen en el organismo durante años o incluso décadas, permitiendo respuestas mucho más rápidas y eficaces frente a exposiciones posteriores al mismo microorganismo. Este fenómeno constituye la base inmunológica del éxito de la vacunación.
Además de inducir respuestas inmunitarias, las células dendríticas desempeñan un papel esencial en el mantenimiento de la tolerancia frente a antígenos propios y antígenos ambientales inocuos. En ausencia de señales inflamatorias, pueden presentar antígenos propios sin expresar niveles elevados de moléculas coestimuladoras, induciendo apoptosis, anergia o diferenciación de linfocitos T reguladores. Este mecanismo previene el desarrollo de enfermedades autoinmunes y limita respuestas inmunitarias innecesarias frente a proteínas alimentarias o microbiota comensal.
Las células dendríticas constituyen el nexo central entre inmunidad innata y adaptativa porque integran múltiples funciones secuenciales: reconocen microorganismos mediante receptores de inmunidad innata; captan y procesan proteínas antigénicas; cargan péptidos sobre moléculas del complejo principal de histocompatibilidad clase I y clase II; experimentan un proceso de maduración que incrementa su capacidad estimuladora; migran desde los tejidos periféricos hacia los órganos linfoides secundarios; establecen sinapsis inmunológicas con linfocitos T vírgenes; proporcionan señales antigénicas, coestimuladoras y citocínicas indispensables para su activación; inducen la diferenciación de linfocitos T citotóxicos y linfocitos T cooperadores; promueven la formación de memoria inmunológica y mantienen la tolerancia frente a antígenos propios. Gracias a esta extraordinaria integración funcional, las células dendríticas transforman la detección inespecífica de un agente infeccioso en una respuesta adaptativa altamente específica, eficaz y duradera.


Fuente y lecturas recomendadas:
- Abbas, A. K., Lichtman, A. H., & Pillai, S. (2022). Cellular and Molecular Immunology (10th ed.). Elsevier.
- Banchereau, J., & Steinman, R. M. (1998). Dendritic cells and the control of immunity. Nature, 392(6673), 245-252. https://doi.org/10.1038/32588
- Guermonprez, P., Valladeau, J., Zitvogel, L., Théry, C., & Amigorena, S. (2002). Antigen presentation and T cell stimulation by dendritic cells. Annual Review of Immunology, 20, 621-667. https://doi.org/10.1146/annurev.immunol.20.100301.064828
- Iwasaki, A., & Medzhitov, R. (2015). Control of adaptive immunity by the innate immune system. Nature Immunology, 16(4), 343-353. https://doi.org/10.1038/ni.3123
- Janeway, C. A., Travers, P., Walport, M., & Shlomchik, M. J. (2001). Immunobiology: The Immune System in Health and Disease (5th ed.). Garland Science.
- Merad, M., Sathe, P., Helft, J., Miller, J., & Mortha, A. (2013). The dendritic cell lineage: Ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annual Review of Immunology, 31, 563-604. https://doi.org/10.1146/annurev-immunol-020711-074950
- Murphy, K., & Weaver, C. (2022). Janeway’s Immunobiology (10th ed.). W. W. Norton & Company.
- Steinman, R. M., & Cohn, Z. A. (1973). Identification of a novel cell type in peripheral lymphoid organs of mice. The Journal of Experimental Medicine, 137(5), 1142-1162. https://doi.org/10.1084/jem.137.5.1142
- Steinman, R. M. (2007). Dendritic cells: Understanding immunogenicity. European Journal of Immunology, 37(Suppl. 1), S53-S60. https://doi.org/10.1002/eji.200737400
Aprende administración paso a paso

ADMINISTRACION DESDE CERO

